-

-

-
Models Advantages Disadvantages Cell models PTH/PHH -intracellular environment close
to that in vivo-poor accessibility;
-limited cell numbers;
-lot-to-lot variation;
-genetic manipulation impossibleHepaRG -homogeneous source -license protected, which limits wide application;
-induction of differentiation time-consuming
and complicatedHepAD38 -homogeneous source;
-genetic manipulation possible-intracellular environment quite different
from that in vivoHLCZ01 -homogeneous source;
-genetic manipulation possible-not yet widely used NTCP-HepG2/Huh7 -homogeneous source;
-genetic manipulation possible-intracellular environment quite different
from that in vivoAnimal models Transgenic mice -wide applications;
-simple techniques-no detectable cccDNAs in liver;
-no associated hepatitis disease for mouseHuman liver chimeric mice -cccDNAs detectable in liver -high costs;
-unstable results;
-low cccDNA copy numbers;
-complicated techniquesWoolly monkey -cccDNAs detectable in liver -high costs;
-long time periods for study;
-ethical concernsChimpanzee -cccDNAs detectable in liver;
-situation in vivo similar to that in human-high costs;
-long time periods for study;
-ethical concernsTable 1. A comparison of the advantages and disadvantages for cellular and animal models with detectable cccDNAs
-
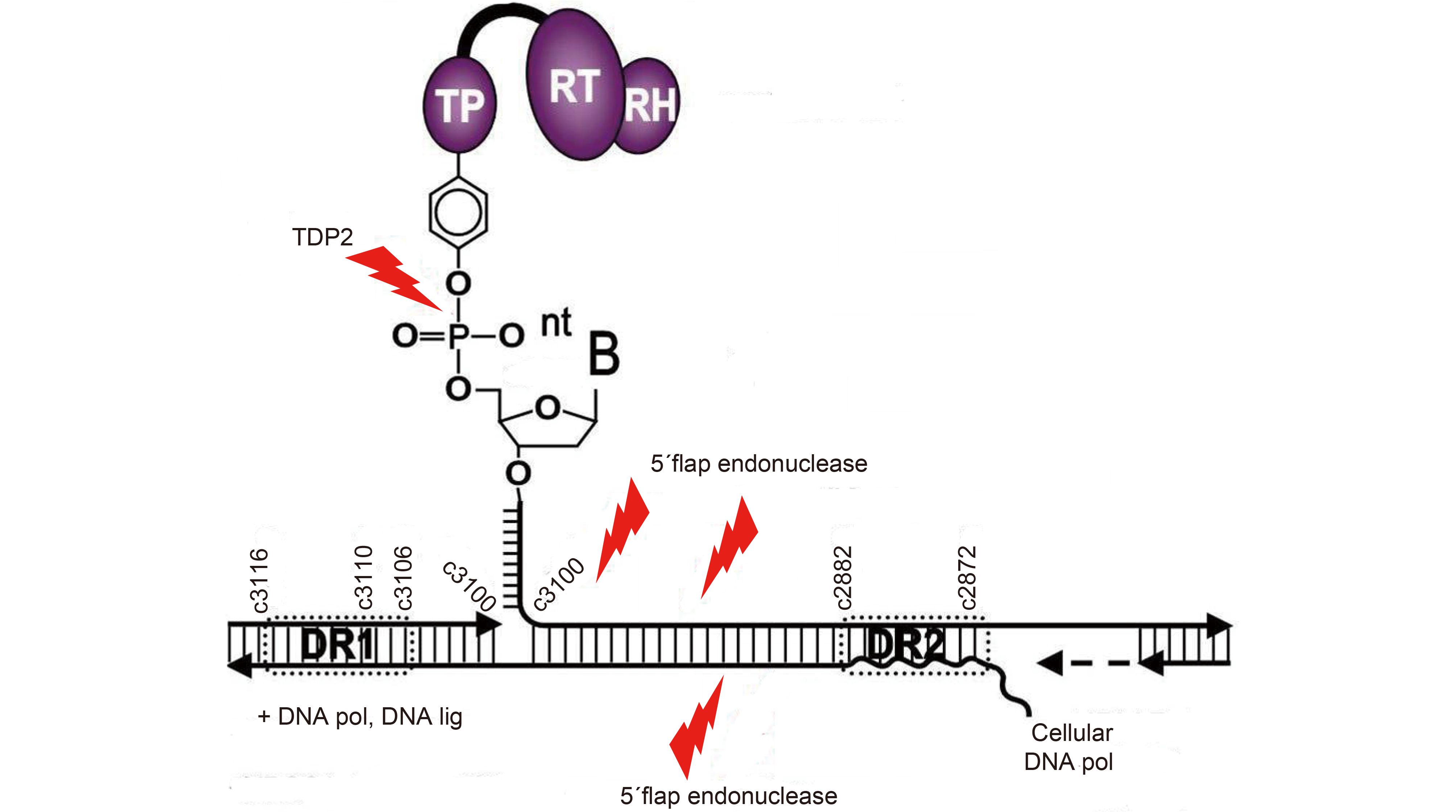
Strategies References Advantages Limitations Future directions Host DNA damage response Pommier et al., 2014;
Königer et al., 2014;
Yang et al., 2014;
Cui et al., 2015;
Qi et al., 2016;
Oqi and Lehmann, 2006;
Oqi et al., 2010;
Ran et al. 2017;
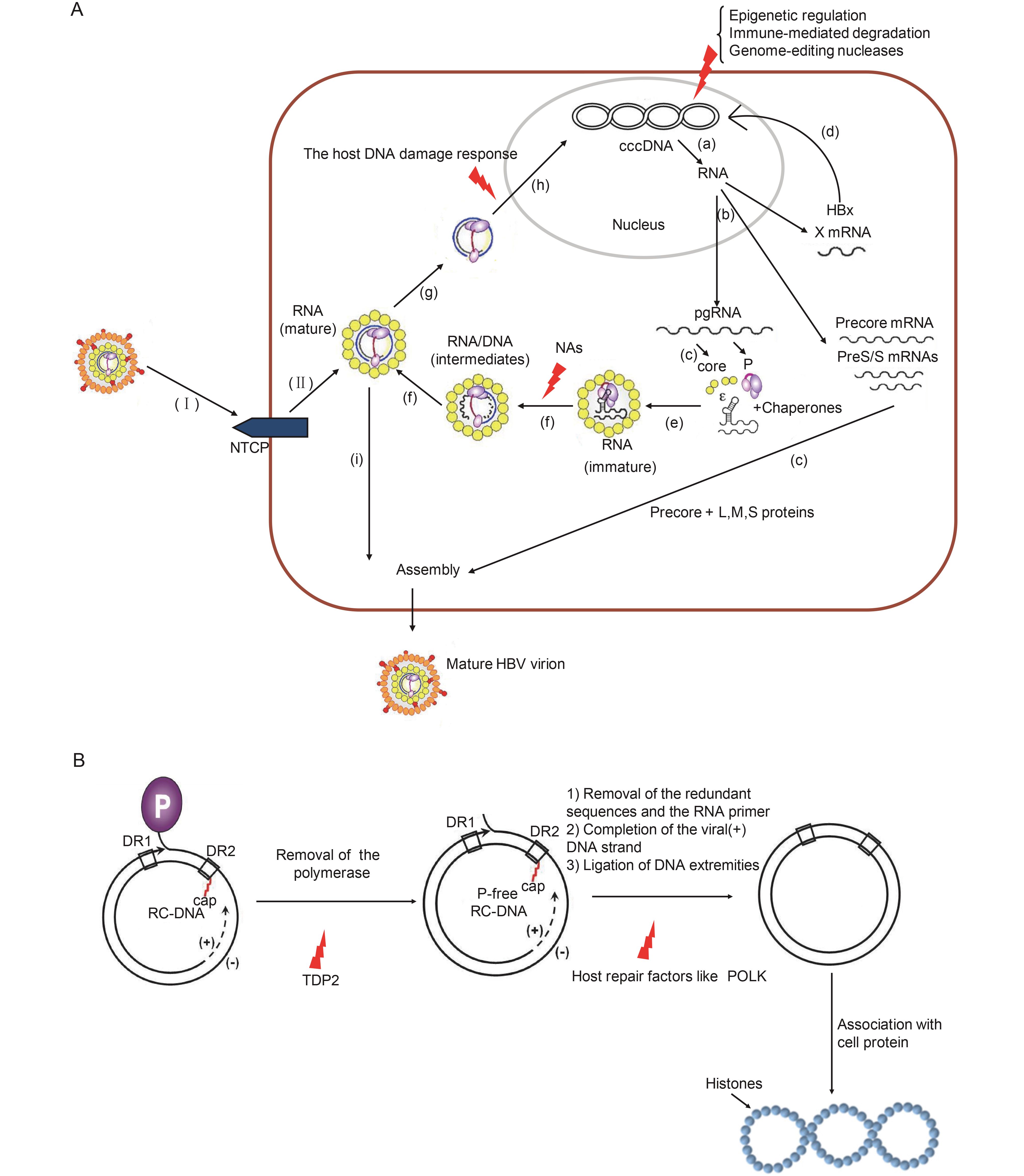
Schreiner and Nassal, 2017.The present experimental evidence shows many members of the host DNA damage response, such as TDP2, DNA polymerase, especially DNA polymerase κ, and the host DNA ligases are involved in the conversion of RC-DNA to cccDNA. Interference with DNA repair response may cause severe genetic instability of host hepatocytes. The study of how the host DNA damage response is involved in the conversion of RC-DNA to cccDNA is just at its beginning. Through knockout of a defined DNA repair gene, an individual host factor in the DNA repair pathway can be investigated for its involvement in cccDNA formation. Epigenetic regulation of cccDNA Belloni et al., 2009;
Guo et al., 2014;
Rivière et al., 2015;
Decorsière et al., 2016;
Murphy et al., 2016;
Niu et al., 2017;
Schubeler, 2015;
Vivekanandan et al., 2010;
Hong et al., 2017;
Palumbo et al., 2015.HBx is required for cccDNA transcriptional activity; thus, it represents an attractive anti-cccDNA target. DNMTs could be utilized to inhibit cccDNA transcription. IL6 has a repressive effect on HBV replication. The mechanism of HBx-mediated cccDNA transcription activation is to a large extent unclear. cccDNA functional inactivation, instead of elimination, may be only a transient solution. The role of DNMTs in HBV DNA methylation has not been clearly identified. Epigenetic regulation of cccDNA transcription activity should be addressed in detail, including post-translational modifications of histones, nucleosome spacing, non-coding RNAs, etc. Immune-mediated degradation Lucifora et al., 2014;
Luo et al., 2016;
Kitamura et al., 2013.High doses of IFN-α or LTβR-agonists were able to trigger noncytolytic degradation of cccDNA. This underscores the value of activating the innate immune response. A fraction of the cccDNA pool is refractory to further reduction. Studies, such as of the Toll-like receptor 7 agonist GS-9620, are indicative of the importance of reactivating host innate response. Genome-editing nucleases Seeger and Sohn, 2016;
Lin et al., 2014;
Ramanan et al., 2015;
Zhen et al., 2015;
Lin et al., 2015;
Fu et al., 2013.The application of artificial endonucleases has promoted cccDNA-related studies. More than 90% of HBV DNAs were cleaved by Cas9. The CRISPR/Cas9 system has been used to target HBV DNA, including cccDNA in cell culture and in the murine model. It is so far unconfirmed that the nuclease has been delivered to each persistent infected hepatocyte. The potential off-target effects may have an adverse effect on liver cell function, especially in the presence of long-acting effector enzymes. The CRISPR/Cas9 system can lead to indels in the host genomes. Finally, excessive amounts of pre-existing RC-DNA may supplement the cccDNA source. How nucleases enter into all HBV-infected hepatocytes to attack cccDNA, and how to avoid off-target effects of CRISPR/Cas9 etc. remain major challenges. Table 2. Comparison of four strategies against HBV cccDNA
Figure 2 个
Table 2 个