-

Prion Protein Protects Cancer Cells against Endoplasmic Reticulum Stress Induced Apoptosis
2019, 34(2): 222 doi: 10.1007/s12250-019-00107-2
Received: 09 November 2018 Accepted: 28 February 2019 Published: 24 April 2019Unfolded protein response (UPR) is an adaptive reaction for cells to reduce endoplasmic reticulum (ER) stress. In many types of cancers, such as lung cancer and pancreatic cancer, cancer cells may harness ER stress to facilitate their survival and growth. Prion protein (PrP) is a glycosylated cell surface protein that has been shown to be up-regulated in many cancer cells. Since PrP is a protein prone to misfolding, ER stress can result in under-glycosylated PrP, which in turn may activate ER stress. To assess whether ER stress leads to the production of under-glycosylated PrP and whether underglycosylated PrP may contribute to ER stress thus leading to cancer cell apoptosis, we treated different cancer cells with brefeldin A (BFA), thapsigargin (Thps), and tunicamycin (TM). We found that although BFA, Thps, and TM treatment activated UPR, only ATF4 was consistently activated by these reagents, but not other branches of ER stress. However, the canonical PERK-eIF2α-ATF4 did not account for the observed activation of ATF4 in lung cancer cells. In addition, BFA, but neither Thps nor TM, significantly stimulated the expression of cytosolic PrP. Finally, we found that the levels of PrP contributed to anti-apoptosis activity of BFA-induced cancer cell death. Thus, the pathway of BFA-induced persistent ER stress may be targeted for lung and pancreatic cancer treatment. -
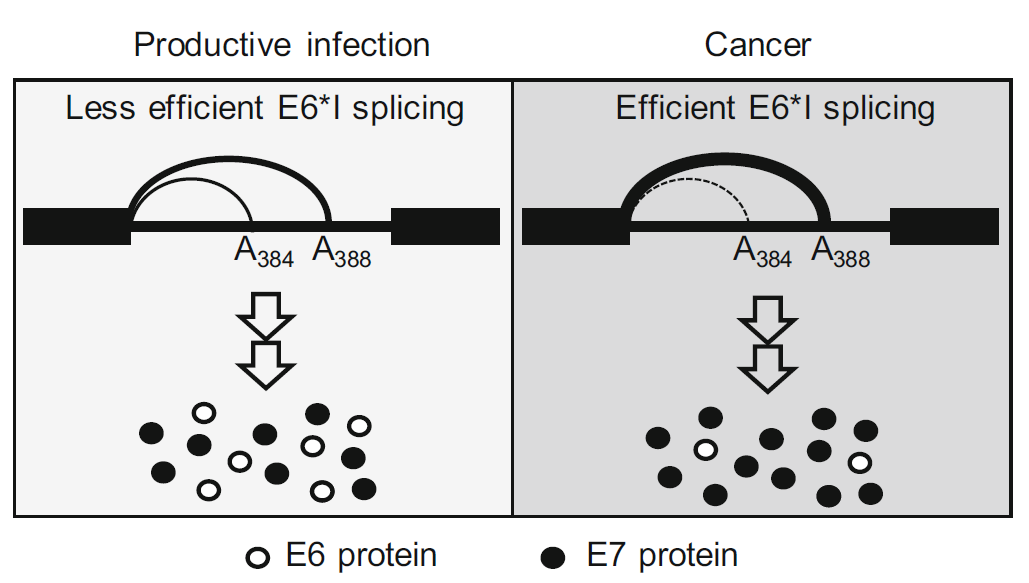
HPV18 Utilizes Two Alternative Branch Sites for E6*I Splicing to Produce E7 Protein
2019, 34(2): 211 doi: 10.1007/s12250-019-00098-0
Received: 17 December 2018 Accepted: 21 February 2019 Published: 03 April 2019Human papillomavirus 18 (HPV18) E6 and E7 oncogenes are transcribed as a single bicistronic E6E7 pre-mRNA. The E6 ORF region in the bicistronic E6E7 pre-mRNA contains an intron. Splicing of this intron disrupts the E6 ORF integrity and produces a spliced E6*I RNA for efficient E7 translation. Here we report that the E6 intron has two overlapped branch point sequences (BPS) upstream of its 3' splice site, with an identical heptamer AACUAAC, for E6*I splicing. One heptamer has a branch site adenosine (underlined) at nt 384 and the other at nt 388. E6*I splicing efficiency correlates to the expression level of E6 and E7 proteins and depends on the selection of which branch site. In general, E6*I splicing prefers the 3'ss-proximal branch site at nt 388 over the distal branch site at nt 384. Inactivation of the nt 388 branch site was found to activate a cryptic acceptor site at nt 636 for aberrant RNA splicing. Together, these data suggest that HPV18 modulates its production ratio of E6 and E7 proteins by alternative selection of the two mapped branch sites for the E6*I splicing, which could be beneficial in its productive or oncogenic infection according to the host cell environment. -

MicroRNA-135a Modulates Hepatitis C Virus Genome Replication through Downregulation of Host Antiviral Factors
2019, 34(2): 197 doi: 10.1007/s12250-018-0055-9
Received: 24 July 2018 Accepted: 13 September 2018 Published: 09 November 2018Cellular microRNAs (miRNAs) have been shown to modulate HCV infection via directly acting on the viral genome or indirectly through targeting the virus-associated host factors. Recently we generated a comprehensive map of HCV– miRNA interactions through genome-wide miRNA functional screens and transcriptomics analyses. Many previously unappreciated cellular miRNAs were identified to be involved in HCV infection, including miR-135a, a human cancerrelated miRNA. In the present study, we investigated the role of miR-135a in regulating HCV life cycle and showed that it preferentially enhances viral genome replication. Bioinformatics-based integrative analyses and subsequent functional assays revealed three antiviral host factors, including receptor interacting serine/threonine kinase 2 (RIPK2), myeloid differentiation primary response 88 (MYD88), and C-X-C motif chemokine ligand 12 (CXCL12), as bona fide targets of miR-135a. These genes have been shown to inhibit HCV infection at the RNA replication stage. Our data demonstrated that repression of key host restriction factors mediated the proviral effect of miR-135a on HCV propagation. In addition, miR-135a hepatic abundance is upregulated by HCV infection in both cultured hepatocytes and human liver, likely mediating a more favorable environment for viral replication and possibly contributing to HCV-induced liver malignancy. These results provide novel insights into HCV–host interactions and unveil molecular pathways linking miRNA biology to HCV pathogenesis. -

Manipulating the Interferon Signaling Pathway: Implications for HIV Infection
2019, 34(2): 192 doi: 10.1007/s12250-019-00085-5
Received: 29 September 2018 Accepted: 14 December 2018 Published: 14 February 2019During human immunodeficiency virus (HIV) infection, type Ⅰ interferon (IFN-Ⅰ) signaling induces an antiviral state that includes the production of restriction factors that inhibit virus replication, thereby limiting the infection. As seen in other viral infections, type Ⅰ IFN can also increase systemic immune activation which, in HIV disease, is one of the strongest predictors of disease progression to acquired immune deficiency syndrome (AIDS) and non-AIDS morbidity and mortality. Moreover, IFN-Ⅰ is associated with CD4 T cell depletion and attenuation of antigen-specific T cell responses. Therefore, therapeutic manipulation of IFN-Ⅰ signaling to improve HIV disease outcome is a source of much interest and debate in the field. Recent studies have highlighted the importance of timing (acute vs. chronic infection) and have suggested that specific targeting of type Ⅰ IFNs and their subtypes may help harness the beneficial roles of the IFN-Ⅰ system while avoiding its deleterious activities. -
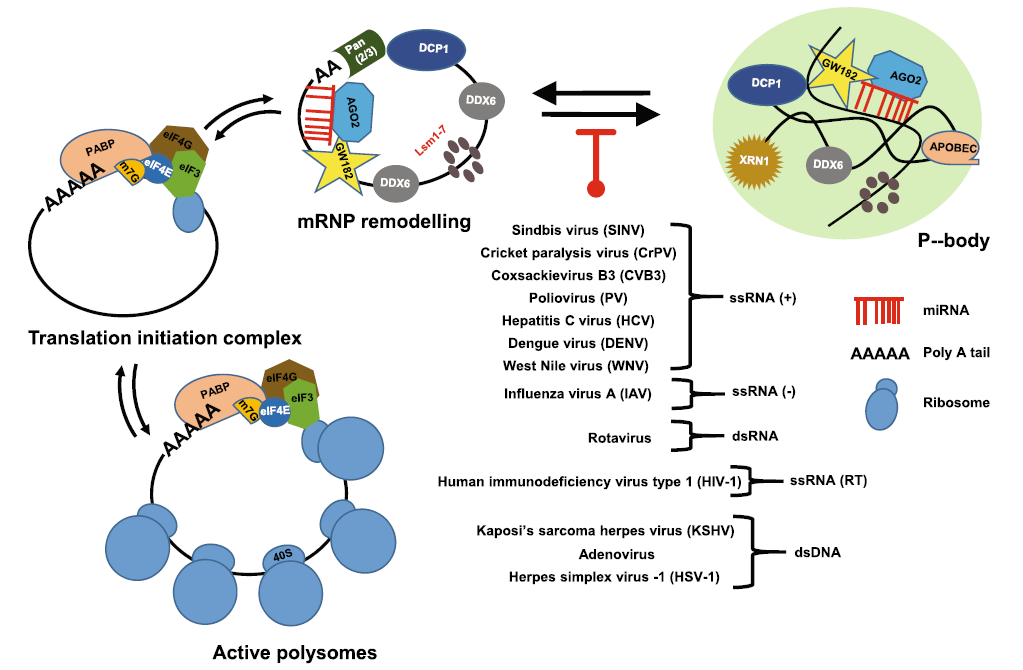
Viral Regulation of RNA Granules in Infected Cells
2019, 34(2): 175 doi: 10.1007/s12250-019-00122-3
Received: 15 January 2019 Accepted: 02 April 2019 Published: 29 April 2019RNA granules are cytoplasmic, microscopically visible, non-membrane ribo-nucleoprotein structures and are important posttranscriptional regulators in gene expression by controlling RNA translation and stability. TIA/G3BP/PABP-specific stress granules (SG) and GW182/DCP-specific RNA processing bodies (PB) are two major distinguishable RNA granules in somatic cells and contain various ribosomal subunits, translation factors, scaffold proteins, RNA-binding proteins, RNA decay enzymes and helicases to exclude mRNAs from the cellular active translational pool. Although SG formation is inducible due to cellular stress, PB exist physiologically in every cell. Both RNA granules are important components of the host antiviral defense. Virus infection imposes stress on host cells and thus induces SG formation. However, both RNA and DNA viruses must confront the hostile environment of host innate immunity and apply various strategies to block the formation of SG and PB for their effective infection and multiplication. This review summarizes the current research development in the field and the mechanisms of how individual viruses suppress the formation of host SG and PB for virus production. -
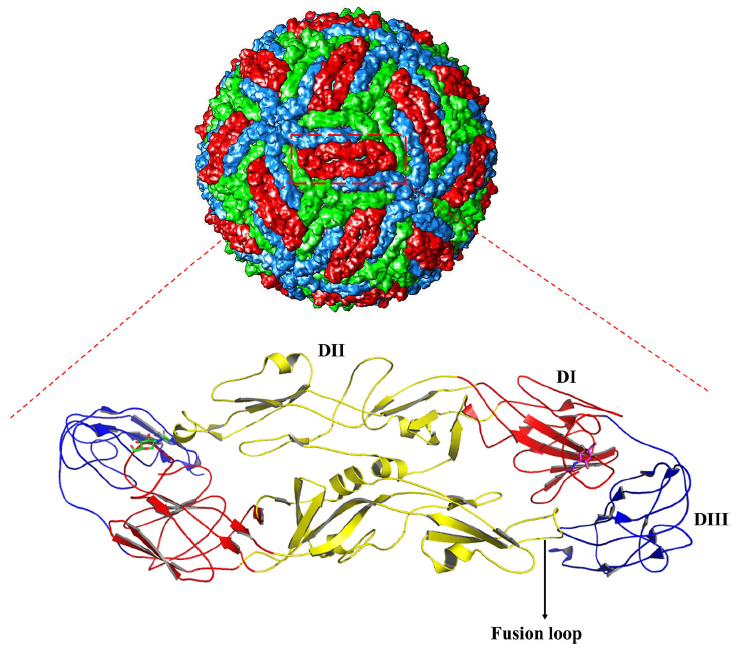
Development of Neutralizing Antibodies against Zika Virus Based on Its Envelope Protein Structure
2019, 34(2): 168 doi: 10.1007/s12250-019-00093-5
Received: 25 October 2018 Accepted: 28 January 2019 Published: 24 April 2019As we know more about Zika virus (ZIKV), as well as its linkage to birth defects (microcephaly) and autoimmune neurological syndromes, we realize the importance of developing an efficient vaccine against it. Zika virus disease has affected many countries and is becoming a major public health concern. To deal with the infection of ZIKV, plenty of experiments have been done on selection of neutralizing antibodies that can target the envelope (E) protein on the surface of the virion. However, the existence of antibody-dependent enhancement (ADE) effect might limit the use of them as therapeutic candidates. In this review, we classify the neutralizing antibodies against ZIKV based on the epitopes and summarize the resolved structural information on antibody/antigen complex from X-ray crystallography and cryo-electron microscopy (cryo-EM), which might be useful for further development of potent neutralizing antibodies and vaccines toward clinical use. -

Flu DRiPs in MHC Class Ⅰ Immunosurveillance
2019, 34(2): 162 doi: 10.1007/s12250-018-0061-y
Received: 28 August 2018 Accepted: 30 September 2018 Published: 19 November 2018Since the publication of the DRiP (defective ribosomal product) hypothesis in 1996, numerous studies have addressed the contribution of DRiPs to generating viral antigenic peptides for CD8+ T cell immunosurveillance. Here, we review studies characterizing the generation of antigenic peptides from influenza A virus encoded DRiPs, discuss the many remaining mysteries regarding the nature of their co-translational generation, and speculate on where the future might lead. -
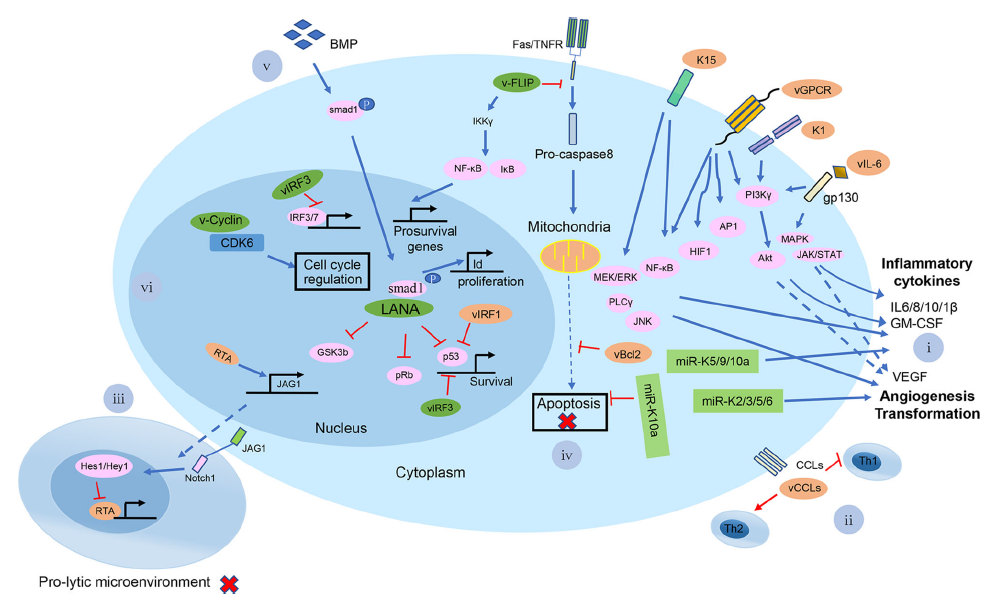
Towards Better Understanding of KSHV Life Cycle: from Transcription and Posttranscriptional Regulations to Pathogenesis
2019, 34(2): 135 doi: 10.1007/s12250-019-00114-3
Received: 29 November 2018 Accepted: 14 March 2019 Published: 25 April 2019Kaposi's sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus-8 (HHV-8), is etiologically linked to the development of Kaposi's sarcoma, primary effusion lymphoma, and multicentric Castleman's disease. These malignancies often occur in immunosuppressed individuals, making KSHV infection-associated diseases an increasing global health concern with persistence of the AIDS epidemic. KSHV exhibits biphasic life cycles between latent and lytic infection and extensive transcriptional and posttranscriptional regulation of gene expression. As a member of the herpesvirus family, KSHV has evolved many strategies to evade the host immune response, which help the virus establish a successful lifelong infection. In this review, we summarize the current research status on the biology of latent and lytic viral infection, the regulation of viral life cycles and the related pathogenesis. -
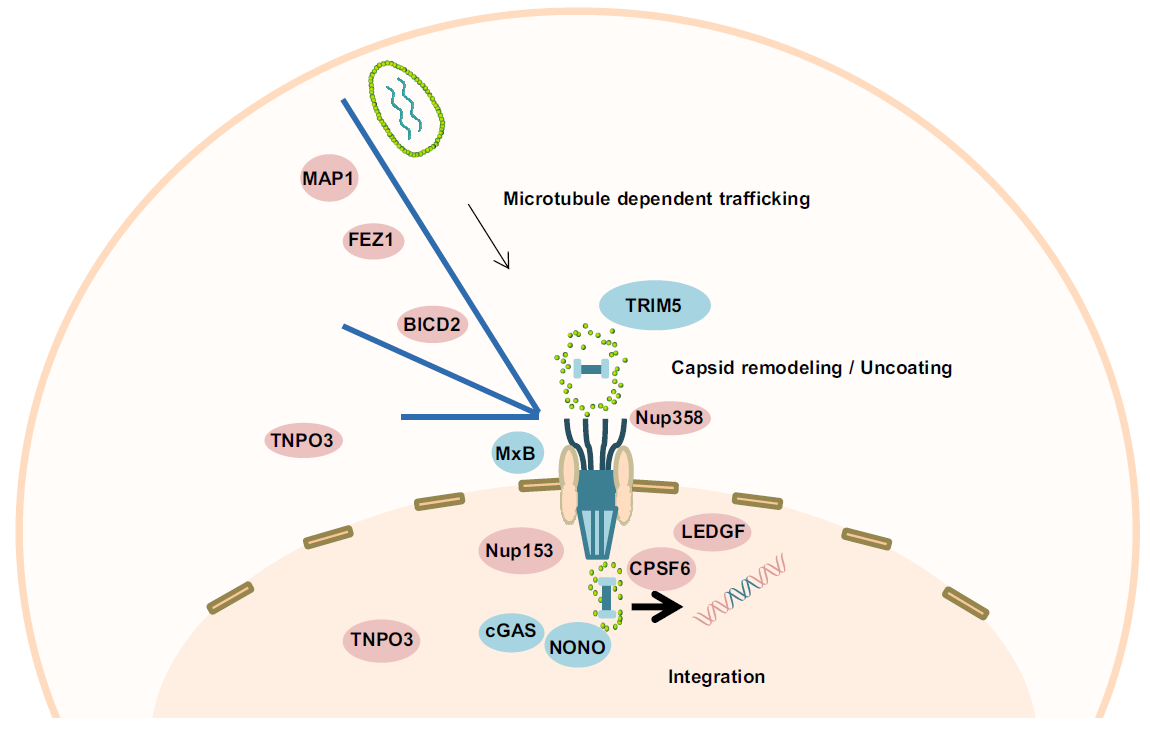
Multiple Roles of HIV-1 Capsid during the Virus Replication Cycle
2019, 34(2): 119 doi: 10.1007/s12250-019-00095-3
Received: 17 November 2018 Accepted: 16 January 2019 Published: 26 April 2019Human immunodeficiency virus-1 capsid (HIV-1 CA) is involved in different stages of the viral replication cycle. During virion assembly, CA drives the formation of the hexameric lattice in immature viral particles, while in mature virions CA monomers assemble in cone-shaped cores surrounding the viral RNA genome and associated proteins. In addition to its functions in late stages of the viral replication cycle, CA plays key roles in a number of processes during early phases of HIV-1 infection including trafficking, uncoating, recognition by host cellular proteins and nuclear import of the viral preintegration complex. As a result of efficient cooperation of CA with other viral and cellular proteins, integration of the viral genetic material into the host genome, which is an essential step for productive viral infection, successfully occurs. In this review, we will summarize available data on CA functions in HIV-1 replication, describing in detail its roles in late and early phases of the viral replication cycle. -