









HTML
-
Foamy viruses (FVs) are members of Retroviridae. They represent a distinct family of complex retroviruses and have been isolated from a number of species including primates (Achong B G, et al., 1971; Bieniasz P D, et al., 1995; Broussard S R, et al., 1997), bovines (Johnson R H, et al., 1983), felines (Mochizuki M, et al., 1990), and equines (Tobaly-Tapiero J, et al., 2000). Although FVs cause substantial cytopathic effects (CPEs) in tissue culture, they only result in persistent infections in their hosts and have not been linked to any known disease (Linial M L, 1999).
Similar to other retroviruses, all FVs characterized to date possess genomic structures with the classical gag, pol, and env structural genes and additional open reading frames (ORFs) that are under the control of the 5′-long terminal repeat (LTR) and an internal promoter (IP) located at the 3′-end of the env gene. However, there are also some distinct features found in FVs, such as the length of genome and characteristics of pol mRNA. FV genomes range from 11, 956-to 13, 246-kb in length and are the largest in the retrovirus family. FV LTRs are also longer than those found in other retroviruses, except for those in mouse mammary tumor virus (MMTV) or in human immunodeficiency virus type 1 (HIV-1). In addition, the pol gene encodes a pol-specific mRNA, and the pol gene products are not generated by frame-shifting or by the termination suppression mechanism (Enssle J, et al., 1996; Konvalinka J, et al., 1995).
Most research has focused on primate FVs, and very little is known about non-primate FVs, including bovine foamy virus (BFV). BFV infection is widespread in farm animals around the world (Appleby R C, 1979; Bouillant A M, et al., 1982; Lucas M H, et al., 1986). Although BFV has not been directly linked to any bovine diseases in vivo, infection of bovine cells in culture frequently leads to CPE and formation of syncytium (Pamba R, et al., 1999). Concomitant infection by BFV, bovine leukaemia virus (BLV), and/or bovine immunodeficiency virus (BIV) has been documented (Amborski G F, et al., 1989; Jacobs R M, et al., 1995; Jacobs R M, et al., 1992). So far, there are four full-length BFV genome sequences in GenBank: BFV (USA, GenBank: U94514), BFV100 (Poland, GenBank: JX307861.1), BFV Riems (Germany, GenBank: JX307862.1) and BFV3026 (China, GenBank: AY134750). Here, we report a full-length infectious genomic DNA clone of BFV3026 and show its ability to establish infection in fetal canine thymus cells (Cf2Th) and to produce CPE in vitro. In addition, we demonstrate the presence of intra-and extracellular BFV3026 particles in Cf2Th cultures.
-
The fetal canine thymus cell line (Cf2Th) was kindly provided by Prof. Jin-Ming GAO (Peking Union Medical College). Primary fetal bovine lung cells (FBLs) were isolated in our lab (Liu S, et al., 1997). Cf2Th and FBL cells were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% (v/v) fetal calf serum, 105 U/L penicillin, and 0.1 g/L streptomycin at 37 ℃ in 5% CO2. BFV3026 was stored in our lab and cultured with Cf2Th cells.
-
Cf2Th cells (5×106) infected with BFV3026 for 2 days were harvested, washed with 10 mL buffer K (20 mmol/L HEPES, 140 mmol/L KCl, 5 mmol/L MgCl2, 1 mmol/L DL-Dithiothreitol [DTT]), and lysed with 250 μL buffer K and 7.5 μL 0.5% Triton X-100 for 10 minutes at room temperature. After centrifugation, the pellet was dissolved in 400 μL TE and 90 μL 5 mol/L NaCl at 4 ℃ overnight. The supernatant after centrifugation was extracted with phenol, chloroform:isoamylalcohol (24:1), and then precipitated for DNA in ethanol with 0.3 mol/L NaAc at -20 ℃ for 1 hour. The DNA pellet was then washed with 70% ethanol, dissolved in 20 μL TE, and stored at -20 ℃.
-
BFV3026 Hirt DNA was used as a template for BFV3026 genome amplification, the genome DNA was subcloned into a pBluescript SK (-) vector. A 4.0-kb fragment comprising of 5'-LTR and part of gag gene was amplified by EX Taq polymerase (TaKaRa) with primer P1 (5'-AATAAGCGGCCGCTGTGGTGGGAAGAC-3') and P2 (5'-ACCCCAAGAATTCCACTGTCTCCTTTGC-3') and digested by Not Ⅰ and EcoR Ⅰ, then ligated into pBluescript SK (-) with the same sticky end, named pBS-S1. The other 8.0-kb fragment containing pol, env and 3'-LTR was amplified with primer P3 (5'-AGACAGTGGAATTCTTGGGGTTTTC-3') and P4 (5'-ACTGGTACCTGAGATTGTTGTGACCTTC-3') and digested by EcoR Ⅰ and Kpn Ⅰ, then ligated into pBS-S1 to generate the full length clone, named pBS-BFV.
-
Analysis of BFV3026 Tas (BTas), Bet (BBet), and Gag expression was performed as described previously (Wang J, et al., 2010). Briefly, Cf2Th cells were transfected with pBS or pBS-BFV or infected by BFV3026. After 48 hours, the cells were washed with phosphate-buffered saline (PBS) twice and lysed, boiled, and separated by 12% polyacrylamide gel electrophoresis, then analysed with the indicated antibodies.
-
Total RNA was isolated from the transfected or infected cells with TRIzol reagent (Life Technologies/Invitrogen) and treated with 2 U RQ1 RNase-free DNase (Promega). Genomic DNA was extracted by DNeasy Blood & Tissue kit (QIAGEN). After denaturing the total RNA from plasma at 90 ℃ for 5 minutes, the first-strand cDNA was synthesised in a buffer (25 μL) containing 200 U M-MLV reverse transcriptase (Promega), 0.5 μg oligo deoxythymidine (dT) primer, 0.2 mmol/L deoxynucleoside triphosphate (dNTP), and 20 U rRNasin (Promega). RT was then performed at 37 ℃ for 60 minutes and terminated by boiling for 10 minutes. The 5' sense and 3' antisense primers used to amplify the IP sequence were 5'-CTTAAAAGACCACTAACTTCTC-3' and 5'-TTCGTGGAGATCCAACCGAAGTAG-3', respectively. Those for the amplification of the gag sequence were 5'-GCGACACGAATTGTGA-3' and 5'-TTCCTTGGCTGCTTCT-3', respectively. PCR amplification was performed in PCR buffer (25 μL) containing the sense and antisense primers (0.4 μmol/L), 0.2 mmol/L dNTP, and 2 U EX Taq polymerase (TaKaRa). The PCR reaction was carried out in a DNA thermal cycler (Bio-Rad) with a program of denaturing at 94 ℃ for 1 minute, annealing at 55 ℃ for 30 seconds, and primer extension at 72 ℃ for 20 seconds. The amplification was carried out for 30 cycles.
-
We collected 6×106 Cf2Th cells 48 hours after transfection or infection in 2 μL fresh medium, froze the cells in liquid nitrogen for 30 seconds, then thawed the cells on ice for 40 minutes. This process was repeated three times. The cell lysate supernatants were filtered with 0.45-μm filters after centrifugation at 5000 rpm for 10 minutes and then were used to infect new BICL or Cf2Th cells.
-
Cf2Th cells (5×104) were seeded on coverslips in a 12-well plate, after 20 hours they were transfected with pBS or pBS-BFV or infected with BFV3026. After 48 hours, the cells were washed with PBS twice followed by fixation in 4% formaldehyde and permeabilisation in 1% Triton X-100. Indirect immunofluorescence was carried out with BTas antiserum (1: 500 dilution) and TRITC-conjugated goat anti-goat secondary antibody (DAKO) diluted in antibody solution (3% bovine serum albumin [BSA], 1% Triton X-100, and 0.05% NaN3 in PBS). Cell nuclei were stained with 4, 6-diamidino-2-phenylindole (DAPI), and fluorescence was observed with an Olympus fluorescence microscope. The procedures were the same for BFV3026-infected Cf2Th cells.
-
BFV3026-infected or pBS-BFV transfected cells were fixed with 3% glutaraldehyde in PBS overnight at 4 ℃. The cells were then post-fixed with 1% aqueous osmium tetroxide for 1 hour at room temperature, dehydrated in ethanol, and embedded in SPI-PON 812. Ultrathin sections were collected on 300-mesh copper grids and stained with uranyl acetate and lead citrate. Samples were then examined using a Philips EM400ST transmission electron microscope.
Cells and virus culture
Hirt DNA extraction
Construction of BFV3026 infectious clone
Western blot analysis
Reverse transcription (RT) and polymerase chain reaction (PCR) amplification
Freeze-thaw cycle
Immunofluorescence assay
Electron microscopy
-
We previously isolated a BFV strain (named BFV3026) from peripheral lymphoid cells of a cow (Liu S, et al., 1997), and a full-length DNA clone of the BFV3026 genome was constructed from sub-genomic DNA fragments based on the sequence of the BFV USA strain (Renshaw R W, et al., 1994) (Figure 1). It was reported that there was a unique EcoR Ⅰ site in the BFV USA strain that could be used to cut the full-length proviral genome into two fragments of 4.0-kb and 8.0-kb in length. Therefore, two pairs of primers were synthesized based on the position of this enzyme site to amplify the BFV3026 genome by two separate fragments (Figure 1). We analyzed and sequenced at least six separate clones for these two fragments, and the results showed that they were identical in sequence. Therefore, one clone of each fragment was selected for further cloning. Using these two fragments, the BFV3026 full-length DNA was cloned into plasmid pBluescript SK (-) as described in the methods, and it was termed pBS-BFV. Analysis of the restriction endonuclease sites demonstrated that this full-length clone had similar restriction endonuclease cleavage patterns as the BFV USA isolate (data not shown), suggesting there were substantial similarities between the isolate and the virus construct.
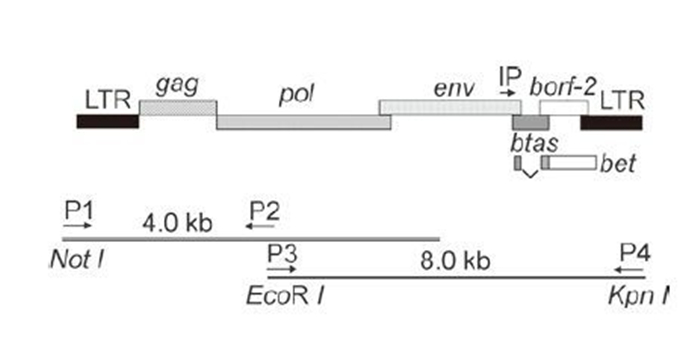
Figure 1. Construction of the BFV3026 infectious clone pBS-BFV. The position of the BFV3026 LTR, the structural genes, accessory genes, and IP are shown with boxes. The unique EcoR Ⅰ restriction endonuclease site and sites for two other endonucleases Not Ⅰ and Kpn Ⅰ in the primers were used in pBS-BFV construction. The positions of the two pairs of primer used for amplification of full-length viral genome are shown.
-
It has been found previously that Cf2Th cells were the most susceptible cells to BFV infection in our lab (data not shown) and were therefore selected for our BFV study. To determine whether pBS-BFV was able to initiate a typical BFV3026 replication cycle and to direct the synthesis of infectious virions, pBS-BFV was transfected into Cf2Th cells and monitored daily for the formation of syncytium by light microscopy. At 2 days post-transfection, typical CPE were observed, and cells that were transfected with empty vector did not show any CPE (Figure 2A). To analyse whether there were viral transcripts, total RNA of the transfected cells was extracted for RT-PCR, and the BFV3026-specific IP sequence was detected in Cf2Th cells transfected with pBS-BFV (Figure 2B). Western blot analyses were carried out to confirm the infectivity of the full-length molecular clone and to determine whether the clone could encode viral proteins. The results showed that the BFV structural protein Gag and accessory proteins, Bet and BTas, were expressed in Cf2Th cells after transfection with pBS-BFV using specific antiserums. The profile of viral proteins expressed in DNA-transfected cells was similar to that observed in BFV3026-infected cells (Figure 2C). Together, these results suggested that the pBS-BFV clone was able to express viral RNA, proteins, and possibly virions.
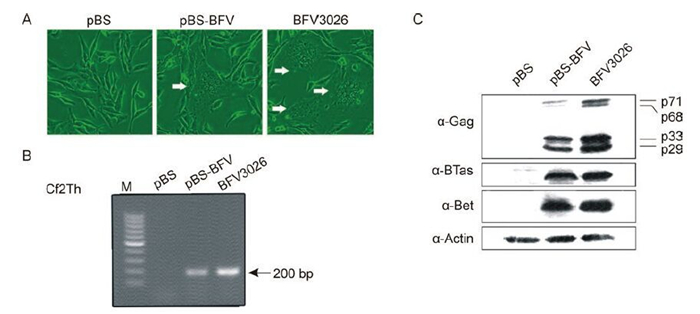
Figure 2. Determination of the infectivity of the full-length BFV3026 genomic clone. (A) Cf2Th cells were transfected with pBS or pBS-BFV or infected with BFV3026. Forty-eight hours later, syncytium could be detected both in pBS-BFV-transfected cells or BFV3026-infected cells (white arrows). (B) Cf2Th cells were transfected with pBS or pBS-BFV or infected with BFV3026. Two days later, total RNA was extracted, and viral transcripts were detected by RT-PCR using IP sequence-specific primers. (C) Cf2Th cells were transfected with pBS or pBS-BFV or infected with wild-type BFV3026. Two days later, Cf2Th cells were harvested and subjected to western blot assays using the indicated antibodies.
To demonstrate the specificity of the CPE, pBS and pBS-BFV-transfected Cf2Th cells were subjected to immunofluorescence assay using BTas antiserum. pBS-BFV-transfected Cf2Th cells showed bright staining with the BTas antiserum similar to infected cells, but Cf2Th cells transfected with pBS vector did not exhibit immunofluorescence (Figure 3). In addition, we performed electron microscopy to determine if virus particles were present. When ultrathin sections of pBS-BFV-transfected cells were observed under an electron microscope, abundant virus particles were seen inside and outside of Cf2Th cells (Figure 4). These particles were 100–200 nm in diameter and had a clear central core that was surrounded by multiple spikes, which is in accordance with the typical appearance of FVs.
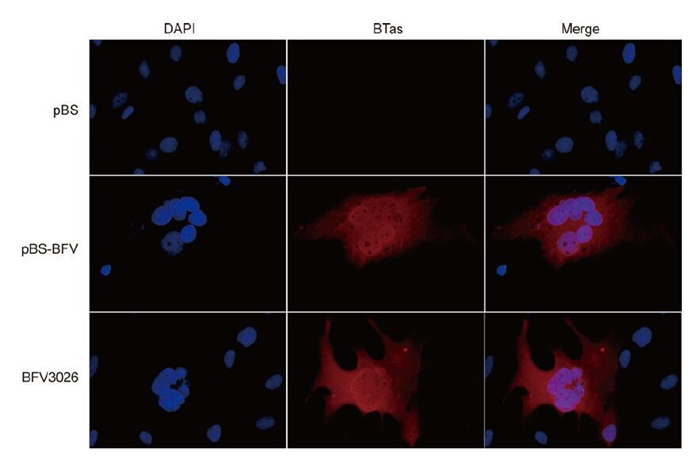
Figure 3. Immunofluorescence labelling of BFV3026-infected and pBS-BFV-transfected Cf2Th cells. Cf2Th cells were transfected with pBS or pBS-BFV or infected with BFV3026, and 2 days later, the Cf2Th cells were labelled with BTas antiserum (dilution 1: 500) and DAPI (dilution 1: 5000).
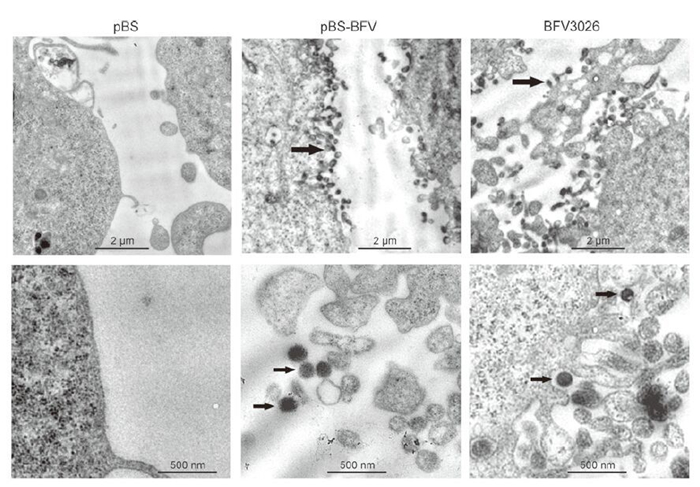
Figure 4. Electron microscopy of ultrathin sections of pBS-BFV-transfected cells. Electron micrographs of cells transfected with pBS, pBS-BFV, or infected with BFV3026 virus at two different magnifications, as indicated. The virus particles exhibited typical FV appearances with enveloped particles surrounded by spikes and clear central cores (black arrows).
To further determine whether infectious viral particles could be generated, lysates from the transfected cells were tested for their infective ability. Cf2Th cells were transfected with pBS or pBS-BFV or infected with BFV3026. Two days later, the cells were harvested and lysed after three freeze-thaw cycles as described in the methods, then BICL and Cf2Th cells were infected with these cell lysates. After three to five parallel passages, some BICL cells turned to green, indicating that they were infected by BFV3026 from Cf2Th cell lysates transfected with pBS-BFV or infected BFV3026, but not the negative control (Figure 5A). Meanwhile, the total RNA and genomic DNA of infected Cf2Th cells were extracted, and the BFV3026 IP sequence was detected in cells infected with pBS-BFV-transfected or BFV3026-infected Cf2Th cell lysates, which was similar to BFV3026 protein expression (Figure 5B). In addition, FBL cells were infected with pBS-BFV-transfected Cf2Th cell lysates and passaged four times, and the BFV3026-specific gag sequence could be detected using RT-PCR (Figure 5C). These results suggested that lysates from pBS-BFV-transfected cells were able to generate BFV3026 mRNA, proteins, and infectious BFV3026 virions.
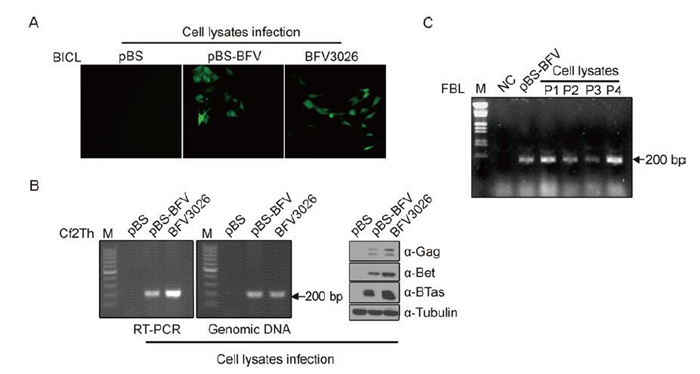
Figure 5. Detection of the infectivity of cell lysates from pBS-BFV-transfected cells. (A) BICLs were infected with lysates from Cf2Th cells transfected with pBS or pBS-BFV or infected with BFV3026. Green fluorescence was detected after five passages. (B) Cf2Th cells were infected with the same amount of cell lysates as in (A), and the viral transcripts, genomic DNA sequence in infected Cf2Th cells, and protein expression were detected by RT-PCR, PCR, and western blotting, respectively. (C) FBL cells were transfected with pBS or pBS-BFV. Two weeks later, the cells were lysed with freeze-thaw methods, and new FBL cells were infected with these cell lysates and passaged for another four generations (P1-4). Viral transcripts were detected by RT-PCR using gag sequence-specific primers. M, DNA molecular weight marker; IP, internal promoter; BICL, BFV3026 indicator cell line; NC, negative control.
-
To our knowledge, only four BFV isolates have been identified to date, and these are from USA, China, Poland, and Germany. After analysing the infectivity of the BFV3026 infectious clone, the full-length BFV3026 genome was sequenced and aligned together with the other three BFV isolates to determine whether genomic organisation was conserved. Sequence analysis revealed that the four virus genomes were mainly conserved with regard to length and organization. BFV structure protein sequences were aligned as shown in Figure 6A, BFVs from USA and China were much more similar and arose from the same branching code. The BFV Gag protein is the smallest of any known FV but has similar overall features. Furthermore, there is a Pro-rich segment of approximately 48 amino acid (AA) residues in size located between the matrix (MA) and capsid (CA) regions, and it contains three Gly–Arg-rich regions instead of the typical zinc finger domain at the C terminus observed in other FVs (Eastman S W, et al., 2001). Comparison of the nucleotide sequence of BFV gag with other FV gag genes showed sequence similarities that ranged from 24% in comparison with simian FV (SFV3) to 32% for equine FV (EFV), which was less than the extent of conservation observed for the pol and env genes. For comparison of BFV3026 and the BFV USA isolate (Table 1), most of the nucleotide substitutions were found to be synonymous changes, only one change led to a substitution in the AA sequence. BFV3026 was more similar to BFV USA than to BFV Riems or BFV 100.
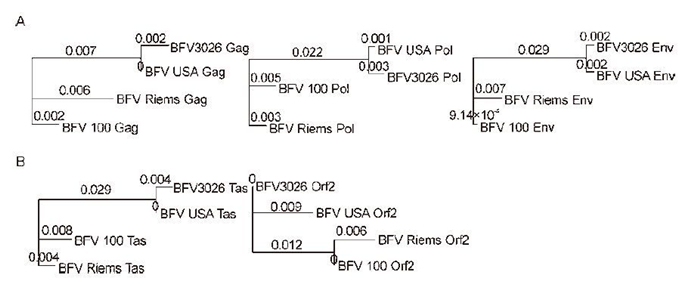
Figure 6. Unrooted phylogenetic tree of complete protein sequences from BFV genomes. Amino acid alignment for subsequent tree analysis was performed using a global alignment with a Blosum62 matrix (Biomatters Geneious, 6.05. Biomatters Ltd., New Zealand). The complete protein sequences of the BFV structure proteins Gag, Pol, and Env were aligned in (A), and auxiliary proteins Tas and Orf2 in (B). A Jukes-Cantor genetic distance model combined with the Neighbor-Joining method and bootstrapping (100 replicates) were used in phylogenetic tree construction. Numbers above the branches show the substitutions per site.
Gag Pol Env AA siteb 3026 USA Riems 100 AA siteb 3026 USA Riems 100 AA siteb 3026 USA Riems 100 160 V · A A 34 I · V V 32 V · · I 182 I · V · 50 N · D D 38 D · E E 189 A · V V 63 I · V V 49 I · M M 199 A V V V 74 A · T T 50 R · K K 377 T · · P 85 V · I I 73 L · F F 434 R · Q · 109 T · A A 115 P · S S 436 C · Y Y 113 E · · G 131 A · V · 501 S · N · 121 Q · · R 174 T · S S 536 S · P P 122 M · T M 182 E · V V 125 K · R · 191 K · E · 156 A · T T 197 L · M · 167 I · V V 272 N D D D 177 I · · V 280 R · K · Tas 210 G R R R 285 K · E E AA siteb 3026 USA Riems 100 326 I · T T 372 R · K K 59 E · V · 329 A T I I 412 D · N N 98 G · D D 458 E K K K 423 K · R R 104 K · · R 484 A · V · 444 P · S S 108 D N N N 537 S · T T 447 L · V V 128 D · N N 574 V · I I 455 Y · H H 148 N · D D 583 M · V V 489 R · K K 151 G · S · 627 R · K K 515 I · A A 152 T · A A 653 I · M M 522 I · A A 171 D · N N 704 A · T T 537 A · G · 216 T · A A 768 T · A A 545 T · S S 222 D · N N 809 V · A A 551 A · D D 827 S · T T 563 K · R R 841 R · K K 583 I · A A 849 I · T T 617 V · I I Orf2 861 S · P P 634 T · A A AA siteb 3026 USA Riems 100 897 Q · · H 641 A T T T 83 W C · · 901 V · I · 662 S N · · 84 P R · · 1029 V A · · 663 K · R R 198 K E · · 1037 S N N N 701 M · I I 225 N · D · 1062 L · S S 731 H · Y Y 244 K · R · 1110 T · · A 735 Y · H H 280 S · P P 1164 G · D D 775 M · V V 287 I · M M 1205 Q · R R 944 A · I · 329 L · P P 1215 V · M M 976 R · G R 331 L · P P 1217 A · T T 984 G E · · "·": Identical with the AA in proteins of BFV3026; a BFV 3026, BFV USA, BFV Riems, and BFV 100 structure and auxiliary proteins were analysed; b Start from the first AA of each protein. Table 1. Amino acid differences in the structure and auxiliary proteins of BFVa
The pol gene encodes the protease (PR), the integrase (IN), and the reverse transcriptase (RT) proteins. The RT-coding region appears to be the most conserved of the regions and has > 60% homology with other FV RT genes. We compared the AA sequence of the Pol protein with the known BFV Pol sequences. BFV3026 possessed five AA substitutions in this region which were found in the conserved motifs of the RT and IN proteins of the BFV USA isolate (Table 1), and more in the BFV isolates from Germany and Poland. Interestingly, in the RT region, the two changes were found in basic residues between the BFV3026 and BFV USA isolates, which were replaced by either a neutral or an acidic AA (Table 1).
We examined the sequence similarity of the BFV env gene with other FVs, and similarity was between 32.9% for SFV-3 and 62% for EFV. We therefore concluded that the Env protein was more conserved than the Gag protein (Holzschu D L, et al., 1998). However, there still were four AA substitutions between BFV3026 and the other BFV sequences in this region (Table 1).
As with all complex retroviruses, BFV encodes several other gene products in addition to the Gag, Pol, and Env proteins. One such gene is btas that encodes the BTas protein. It was previously demonstrated that BTas was a transactivator that could increase virus gene transcription (Wang W, et al., 2010). Another product is the BBet protein, which is generated by alternative splicing. BBet protein shares its N-terminal 35 AAs with BTas, and contains 50 AAs formed after one base pair shift in ORF1 and the entire ORF2 (Figure 1A) (Renshaw R W, et al., 1994). The role of this fusion protein in the virus replication cycle is still unknown (Lecellier C H, et al., 2002; Linial M, 2000). However, Bet has been shown to play a key role in the establishment and maintenance of persistent infection in prototype foamy virus (PFV) (Bock M, et al., 1998).
The sequence identity between BFV3026 accessory proteins and those from other BFV isolates fluctuated from 97% to 99% (Figure 6B), but the homology with other FVs was quite low (Kehl T, et al., 2013). A summary of differences in the BTas and Borf-2 proteins of BFV isolates are shown in Table 1. There were a number of alterations that could lead to potential structural changes. However, the signal-processing program revealed no major structural or functional differences among these BFV isolates.
Construction of the BFV3026 infectious clone
Infectivity of the BFV3026 full-length genomic clone
Polygenetic relationships among BFV isolates
-
In this study, a full-length infectious DNA clone of BFV3026 isolate was constructed. This clone induced the formation of syncytium in transfected Cf2Th cells and generated viral RNA, proteins and virions. In addition, the newly generated infectious particles could be passaged in culture. The sequence alignment of different BFV isolates revealed that the BFV3026 sequence was highly conserved compared with the other three BFVs. We did not successfully amplify viral RNA or detect viral proteins in the supernatant, suggesting that unlike PFV or FFV, BFV cannot release viral particles into the supernatant. We found that freeze-thaw method reduced the infectivity of viral particles from pBS-BFVtransfected Cf2Th cells compared to co-culture, and that it would be necessary to find another way to obtain more infectious viral particles or another full-length DNA clone with better replication capability.
The availability of such an infectious clone could be a useful tool for researchers to perform molecular analyses and manipulations of the virus genome, such as mutagenesis to determine the role of each viral protein in the replicative cycle. Further investigation of their roles in latency, persistence and lytic infection could be carried out. This full-length DNA clone could be used to construct safe and effective retrovirus vectors for gene transfer and therapy. BFV3026 was found to be unlike other FVs, which are normally highly lytic in vitro; it could establish persistent infection in many cell types, including human cells (Yu H, et al., 2003). This broad host range and the ability to establish non-cytopathic infections could allow BFV to be used as a potential vector for gene treatment and therapy.
BFV may be widespread, but the prevalence of infection worldwide is not known. FVs have not yet been linked to any disease in infected hosts. However, these groups of viruses are extremely cytopathic in infected cells in vitro. This difference in pathogenesis in vitro and in vivo is caused by the host immune response against the virus. Currently, very little is known about the host response to BFV infection. Whether the host immune response is responsible for restraining the virus infection or for suppressing CPEs induced by BFV infection in vivo remains to be elucidated. The availability of an infectious clone as reported here is likely to provide a much-needed tool that can be used in these studies.
-
We thank Maxine L. Linial (University of Washington, USA) for her advice on this project and discussions regarding to the manuscript and Kai Wu (Princeton University, USA) for editing the paper. This work was supported by grants from the National Natural Science Foundation of China (31070135, 31370182), the Tianjin Research Program of Application Foundation and Advanced Technology (12JCQNJC06100), and New Century Excellent Talents in University (NCET-100508).
-
All the authors declare that they have no competing interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.
-
TJ Bing and H Yu carried out the molecular genetic studies, participated in the sequence alignment, and drafted the manuscript. TJ Bing carried out the western blotting, immunoassays, RT-PCR assays and electron microscopy. L Sun assisted with electron microscopy. TJ Bing, Y Li, J Tan, YQ Geng, and WT Qiao designed the study. All authors read and approved the final manuscript.

 DownLoad:
DownLoad: