









HTML
-
One of the approaches to plant protection against pathogens, particularly viruses, consists of the use of environmentally safe biologically active compounds that induce plant resistance (Edreva A, 2004; Thakur M, et al., 2013). Various polysaccharides have been shown to be promising agents for inducing antiviral resistance in plants: 1, 3;1, 6-β-D-glucans from fungi (Rouhier P, et al., 1995) and brown algae (Reunov A V, et al., 1996), 1, 3;1, 4-β-D-glucan from the lichen Cetraria islandica (Stübler D, et al., 1996), xyloglucan-(Šubikova V, et al., 1994) and galactoglucomannan-derived (Slováková L, et al., 2000) oligosaccharides, κ/β-carrageenan from the red alga Tichocarpus crinitus (Reunov A V, et al., 2004), fucoidan from the brown alga Fucus evanescens (Lapshina L A, et al., 2006; Reunov A V, et al., 2009).
Chitosan, the deacetylation product of chitin, is one of the best-known elicitors of the plant innate immune system (Hadrami A E, et al., 2010). There is a growing interest in its ability to enhance antiviral resistance in plants (Iriti M, et al., 2006; Kulikov S N, et al., 2006; Davydova V N, et al., 2011). However, the mechanisms responsible for the antiviral activity of chitosan are far from being fully elucidated.
According to our hypothesis (Lapshina L A, et al. 2011; Reunov A V, et al., 2013), a significant role in antiviral protection of cells is played by the lytic compartment, in which hydrolases, such as proteases and RNases, can cause the degradation of viral particles. This concept has been supported by a variety of data obtained by other authors. It was shown (Salomon R, 1989) that plants of Ipomoea nil L. infected with sweet potato feathery mottle virus recover from the infection because of the high proteolytic activity induced by the virus. Hatsugai N, et al. (2004) have reported that the vacuolar processing enzyme (a cysteine protease), identified by the authors in tobacco mosaic virus (TMV)-infected Nicotiana plants carrying the N resistance gene, plays a key role in the induction of a virus-mediated hypersensitive response. RNases may be involved in antiviral plant resistance induced by different polysaccharides obtained from marine algae, such as 1, 3;1, 6-β-D-glucan, fucoidan, and κ/β-carrageenan (Lapshina L A, et al., 2011). More specifically, the pathogenesis-related protein 10 from Capsicum annuum has been shown to have antiviral and RNase activities (Park C J, et al., 2004); the relationship between enhanced RNase activity and reduced virus accumulation in transgenic plants has been demonstrated by several authors (Zhang L, et al., 2001; Trifonova E A, et al., 2007); and antiviral proteins from some plants have been reported to possess strong RNase activity (Begam M, et al., 2006; Choudhary N L, et al., 2008). It has been hypothesized (Trifonova E A, et al., 2007) that one of the possible mechanisms for the antiviral effects of apoplastic RNases in plants is the degradation of viral RNAs.
The purpose of this work was to study the effect of chitosan on TMV accumulation, on the activity of hydrolases (proteases, RNases), and on the state of the viral particles in leaves of N. tabacum L. cv. Samsun.
-
Assays were made using 4-week-old tobacco plants N. tabacum L. cv. Samsun grown in a greenhouse. A preparation of the TMV U1 strain, purified according to Otsuki Y, et al. (1977), was used as the inoculum.
-
Chitosan preparation with a molecular weight of 17.0 kDa was obtained from crab shells by hydrolysis with hydrogen peroxide at 37℃ as described previously (Davydova V N, et al., 2000).
-
The leaves from the middle plant tier were detached, cut in half along the main vein and dusted with carborundum. In a series of experiments, the left-hand leaf halves (test) were inoculated with a mixture of TMV (2 μg/mL) and chitosan (1 mg/mL), while the right-hand ones (control) were inoculated with TMV only. In other experiments, leaf halves were rubbed with chitosan (test) or with water (control) 24 h before inoculation with TMV. The treated leaves were washed with water and placed in a humidity chamber that was thentransferred to a constant environment room (24℃, 16 h photoperiod, 16000 lux, 70% relative humidity).
-
To study the effect of chitosan on infectivity in leaves of N. tabacum cv. Samsun 3, 5, and 7 d after inoculation with TMV, two disks, 7 mm in diameter, were cut from 10 test and 10 control leaf halves and kept at -20℃. At the end of each experiment, the disks were homogenized in 0.05 mol/L phosphate buffer, pH 7.0, and the homogenates were inoculated on 10 leaf halves of N. tabacum L. cv. Xanthi-nc. Four days after inoculation, the number of TMV-induced local lesions produced on leaf halves of this host was counted. The relative infectivity in the test samples was determined as the number of local lesions per leaf half of N. tabacum L. cv. Xanthi-nc expressed as a percentage of the control. Experiments were repeated five times.
-
The effect of chitosan on the accumulation of the viral coat protein in leaves of N. tabacum L. cv. Samsun was evaluated in 10 experiments. In each experiment, the preparation of the leaf samples was carried out as described above. The viral coat protein content in the samples was determined by double antibody sandwich enzyme-linked immunosorbent assay (ELISA), using antibodies conjugated with peroxidase as described previously (Reunov A V, et al., 2004).
-
The activities of hydrolases (proteases, RNases) were determined in 50 disks (4 mm in diameter) cut from the leaf halves and were expressed in terms of optical density, as described previously (Lapshina L A, et al., 2011). Experiments were repeated five times.
-
For the electron-microscopic study of TMV particles in suspensions from the infected leaves, leaf pieces were cut with a razor blade into a drop of distilled water. The suspensions obtained were placed on Formvar-coated grids, desiccated and negatively stained for 5 min with 2% phosphotungstic acid (PTA), pH 7.0. The samples were then examined under a LIBRA-120 electron microscope (Carl Zeiss, Germany, Jena). In some experiments, the virus suspensions intended for examination by negative-staining electron microscopy were obtained from leaf tissue pieces fixed for 3 h in 6.5% glutaraldehyde prepared in phosphate buffer, pH 7.4. The diameters of PTA-stained viral particles were measured for 20 randomly selected viewing fields at a total magnification of ×150000. Experiments were repeated three times. In each experiment, the diameters of 400 viral particles from the test sample and 400 particles from the control sample were measured.
In several experiments, we carried out a comparative electron-microscopic study of PTA-stained purified TMV preparations mixed and unmixed with chitosan. ves the SV coat
-
For immuno-electron microscopy examinations, we used the antisera kindly provided by Dr. R V Gnutova and Dr. I I Sybiryakova, collaborators at the Institute of Biology and Soil Science, Russian Academy of Sciences, Vladivostok. The infected leaf fragments were cut with a razor blade in a drop of distilled water, the debris was removed, and a formvar-coated grid stabilized with evaporated carbon film was placed onto the surface of the drop. After 1 min, the grid was dried with a piece of filter paper, transferred onto a drop of TMV-specific antiserum and kept in a humidity chamber for 15 min at 37℃. An antiserum to potato virus X (PVX) was used as control. Samples were thoroughly washed with distilled water, desiccated, stained with 2% PTA, pH 7.0, and examined under the electron microscope.
-
The results of the experiments on the determination of infectivity, the content of viral coat protein, hydrolase activities and the diameter of the virions in the leaves were analyzed statistically using Student's t-test.
Plant and virus material
Preparation of chitosan
Inoculations
Infectivity evaluation
Evaluation of viral coat protein content
Determination of hydrolase activity in leaves
Negative-staining electron microscopy
Immuno-electron microscopy
Statistical analysis of the results
-
In TMV preparations mixed with chitosan, we found that the virions were not infrequently associated with the polysaccharide particles (Figure 1).

Figure 1. Virions of purified TMV preparation, negatively stained with PTA. A: Virions unmixed with chitosan. B: Virions mixed with chitosan. Arrows (B) show chitosan particles, associated with the virions. Scale Bar = 100 nm.
-
Both the infectivity and the viral coat protein content in leaves of N. tabacum L. cv. Samsun inoculated with TMV (2 μg/mL) mixed with chitosan (1 mg/mL) were significantly lower in the early period of infection than in leaves inoculated with TMV only. Thus, 3 d after inoculation, the infectivity (Table 1) and the viral coat protein content (Table 2) in the test leaf tissues were reduced by 63% and 66%, respectively, as compared to the control. Chitosan applied 24 h prior to inoculation with TMV also inhibited the infectivity (Table 1) and the viral coat protein content (Table 2) in the leaves, but to a lesser extent than when TMV and chitosan were inoculated together. As the infection developed, the inhibitory effects of the preparation decreased.
Days after infection Relative infectivity (%), TMV + chitosan Inhibitory effect (%) Relative infectivity (%), TMV after treatment with chitosan for 24 h Inhibitory ffect (%) 3 37 ± 4.1*** 63 59 ± 6.2*** 41 5 59 ± 6.0*** 41 74 ± 7.6** 26 7 74 ± 7.8** 26 82 ± 8.8* 18 TMV concentration, 2 μg /mL; chitosan concentration, 1 mg /mL. The relative infectivity is the number of local lesions per leaf half of N. tabacum L. cv. Xanthi-nc, expressed as a percentage of the untreated control. Each value is the average percentage obtained on 50 leaf halves of N. tabacum L. cv. Xanthi-nc (± standard error). The differences are statistically significant relative to controls at P* < 0.05, P** < 0.01 and P*** < 0.001. Table 1. The effect of chitosan on infectivity in N. tabacum L. cv. Samsun leaves infected with TMV
Days after infection Content of viral coat protein (ng/mm2) Inhibitory effect
(%)Content of viral coat protein (ng/mm2) Inhibitory effect
(%)TMV + chitosan Control TMV after treatment with chitosan for 24 h Control 3 1.5 ± 0.17*** 4.4 ± 0.5 66 2.1 ± 0.23*** 4.1 ± 0.42 49 5 6.1 ± 0.7** 11.5 ± 1.3 47 9.3 ± 0.94* 13.5 ± 1.45 31 7 18 ± 1.8* 25 ± 2.6 28 18.5 ± 1.9 22.3 ± 2.3 17 The viral coat protein content in test and control leaves was measured by ELISA and calculated for leaf disk areas of 1 mm2. Each value is the average of 10 replicates (± standard error). The differences are statistically significant relative to controls at P* < 0.05, P** < 0.01 and P*** < 0.001. Table 2. The effect of chitosan on viral coat protein content in N. tabacum L. cv. Samsun leaves infected with TMV
It should be noted that leaf halves of N. tabacum L. cv. Samsun inoculated with a mixture of TMV and chitosan and incubated in a humidity chamber did not show any toxicity symptoms and they appeared similar to leaf halves inoculated with TMV alone (data not shown).
-
Inoculation of N. tabacum L. cv. Samsun leaves with a mixture of TMV and chitosan resulted in a significant increase in the activities of hydrolytic enzymes (proteases, RNases) in comparison to leaves inoculated with TMV only (Table 3).
Days after infection Hydrolases The activities of hydrolase (OD) TMV + chitosan TMV 3 Proteases 0.53 ± 0.05 0.38 ± 0.04 RNases 0.47 ± 0.05 0.32 ± 0.04 5 Proteases 0.50 ± 0.05 0.35 ± 0.04 RNases 0.38 ± 0.04 0.26 ± 0.03 7 Proteases 0.35 ± 0.04 0.23 ± 0.03 RNases 0.31 ± 0.03 0.21 ± 0.02 The activities of hydrolases were determined in 50 disks (4 mm in diameter) cut from leaf halves and expressed in terms of optical density. Each value is the average of 5 replicates (± standard error). The differences are statistically significant relative to controls at P < 0.05. Table 3. The activity of hydrolases in leaves N. tabacum L. cv. Samsun inoculated with TMV mixed with chitosan and inoculated with TMV alone
-
In electron microscope assays of PTA-stained preparations from the infected leaves, we found that such preparations contained, along with normal TMV particles (18 nm in diameter, 300 nm long), abnormal virions, the diameter and length of which were substantially larger or smaller than the normal values (Figure 2). In PTAstained preparations from infected leaves previously fixed with glutaraldehyde, the ratio between normal and abnormal virions did not differ significantly from the ratio in preparations from leaves that had not been fixed with glutaraldehyde.
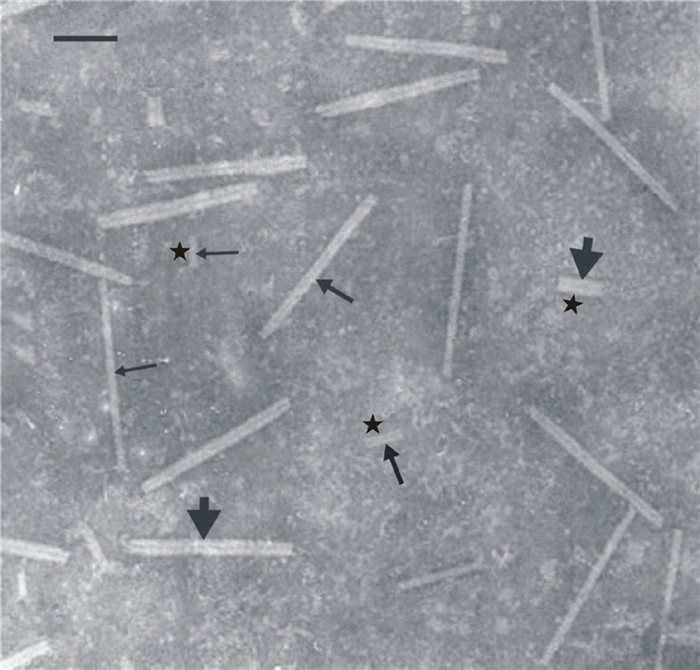
Figure 2. PTA-stained TMV particles from N. tabacum L. cv. Samsun leaf 3 d after inoculation with a mixture of TMV and chitosan. The medium-thickness arrows indicate virions of normal diameter (18 nm); thick arrows indicate swollen virions; thin arrows indicate thin virions. Asterisks show short TMV particles. Scale Bar = 100 nm.
An analysis of the distribution of TMV particle diameters (Table 4) showed that suspensions from the leaves inoculated with a mixture of TMV and chitosan contained a considerably greater number of abnormal virions than those from leaves inoculated with TMV only. Thus, in suspensions from leaves inoculated with a mixture of TMV and chitosan, 26% of viral particles were of normal diameter, 23% were swollen, and 51% were thin. In suspensions from infected untreated leaves, it was found that 63% of virions were of normal diameter, 13% were swollen, and 24% were thin. Calculation of the incidence of short virions showed that in suspensions from leaves infected with a mixture of TMV and chitosan, 40% of the virions were short, whereas in suspensions from control leaves, the incidence was 27%. Amongst the short viral particles, we sometimes observed particles of normal diameter, but much more frequently short particles with diameters greater or smaller than normal were seen.
Diameter of TMV particles, nm Number of TMV particles TMV + chitosan TMV 12 68 ± 7.0** 11 ± 2.5 14 104 ± 12** 22 ± 4.0 16 31 ± 4.0* 65 ± 7.0 18 105 ± 12** 251 ± 25 20 39 ± 5.0** 10 ± 1.5 22 24 ± 3.0 18 ± 2.0 24 29 ± 3.5 23 ± 3.0 The diameters of TMV virions were measured for 20 randomly selected viewing fields at a total magnification of x150000. Experiments were repeated three times. In each experiment, the diameters of 400 viral particles from the test sample and 400 particles from the control sample were measured. Each value is the average of three experiments (± standard error). The differences are statistically significant relative to controls at P* < 0.05 and P** < 0.01. Table 4. Diameter distribution of PTA-stained TMV particles in sap from N. tabacum L. cv. Samsun leaves 3 days after inoculation with TMV mixed with chitosan and with TMV alone
Immuno-electron microscopic studies showed that the virions of normal diameter, as well as the swollen viral particles, were completely decorated with TMV-specific antiserum while the thin virions lost the ability to bind to specific antibodies (Figure 3). In the virions of variable diameter, the antiserim decoration was observed only on the virion parts that were of comparatively large diameter (Figure 3). The antiserum to PVX did not cover the TMV particles.
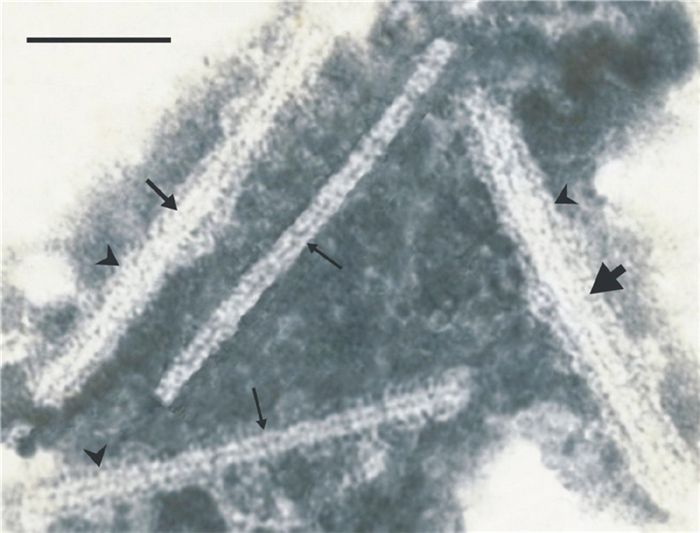
Figure 3. TMV particles from N. tabacum L. cv. Samsun leaf 3 d after inoculation with a mixture of TMV and chitosan. A sap dip from the leaves was exposed to a TMV-specific antiserum and stained with PTA. The medium-thickness, thick, and thin arrows indicate normal, swollen, and thin virions, respectively. The arrowheads show antiserum. The virions of normal diameter, as well as the swollen virions, are completely covered with the antiserum. In the virions of variable diameter (at the bottom of the figure), the antiserum decoration is seen only on the virion parts that are of comparatively large diameter. Scale Bar = 100 nm.
Association of chitosan with TMV particles during mixing
Effect of chitosan on the infectivity and the viral coat protein content in the leaves
Effect of chitosan on the activity of proteases and RNases in the leaves
Influence of chitosan on the state of TMV particles in the leaves
-
The results presented here show that chitosan obtained from crab shells inhibits the development of infection caused by TMV in detached leaves of N. tabacum L. cv. Samsun. This is attested by the preparation-induced decreases in infectivity and viral coat protein content in the leaves. The fact that the highest inhibitory effects of chitosan were observed 3 d after infection of the leaves, and then decreased, indicates that chitosan causes a delay in TMV infection in the early stages. It should be noted that the treatment of the leaves with chitosan and their incubation in a humidity chamber did not result in any toxicity symptoms and the treated leaves appeared similar to control leaves. Hence, the inhibition of TMV infection by chitosan could not be the result of a toxic effect.
In our experiment, the application of chitosan 24 h before virus inoculation noticeably inhibited the accumulation of TMV in the leaves. This effect of the agent seems to be related to its stimulatory action on protective host mechanisms. We also demonstrated that the infectivity and the TMV coat protein content in leaves inoculated with a mixture of the virus and chitosan were substantially lower than in leaves infected 24 h after treatment with chitosan. Therefore, it is reasonable to suppose that the inhibitory effects of chitosan applied together with virus are caused by its influence not only on the host but also on the virus itself.
From electron-microscopic investigations of PTAstained TMV preparations mixed with chitosan, we noted that the polysaccharide particles can be associated with the virions. It seems that association of chitosan with the virions can impede their deproteinization in host cells and thus contribute to inhibition of the infectivity and to a decrease in the viral coat protein content in the leaves. Such a hypothesis had been put forward earlier in relation to the action of a carbohydrate produced by Physarum polycephalum on TMV infection (Mayhew D E, et al., 1971).
The data obtained have demonstrated that suspensions prepared from the infected leaves contain, along with normal TMV particles (18 nm in diameter, 300 nm long), abnormal virions of a diameter and a length significantly larger or smaller than the normal values. The fact that the ratio between normal and abnormal virions in PTAstained suspensions from the infected leaves preliminarily fixed with glutaraldehyde did not differ substantially from the ratio in suspensions from leaves that had not been fixed with glutaraldehyde indicates that the presence of abnormal viral particles in the leaves is not an artifact.
The appearance of abnormal viral particles in leaf cells has been described previously and has been attributed to the development of lytic processes causing destructive changes in virions in vivo (Lapshina L A, et al., 2011; Reunov A V, et al., 2013). We believe that the generation of abnormal virions may occur as follows. Due to a virus-induced change in the environmental conditions (e.g., pH), the viral protein subunits seem to undergo conformational changes resulting in partial loosening and untwining. The loosening of protein subunits probably leads to the appearance of swollen virions, while the partial untwining of the capsid subunit polypeptides leads to limited proteolysis of exposed peptide bonds, resulting in virion thinning. Earlier, Everitt E, et al. (1988) accounted for proteolysis of Adenovirus 2 protein in a similar fashion. The fact that thin TMV particles, unlike the virions of normal diameter, lose the ability to bind to specific antiserum is presumably caused by the elimination, during partial proteolysis of the capsid subunits, of determinants responsible for binding to antibodies.
The presence in the suspensions of TMV particles that are longer than 300 nm can be explained by the ability of virions to link end-to-end with each other, which has been noted previously (Esau K, 1968). The presence of short TMV particles of normal diameter observed sometimes in the suspensions can probably be attributed to incompleteness of assembly. The appearance of short TMV particles of a diameter greater or smaller than that of normal particles can presumably be explained by the fact that, when the stability of the protein virion coat is disturbed, viral RNA becomes available for RNase attack, resulting in the fragmentation (cross-cutting) of virions. The data obtained demonstrate that chitosan treatment of the leaves promotes the formation of such fragmented virions.
The data presented on the chitosan-mediated increase in the number of thin and fragmented viral particles in infected leaves suggest the existence of a correlation between the stimulation of hydrolase (protease, RNase) activities and the intracellular destruction of virions. We believe that the increase in protease and RNase activities, which facilitates destruction of viral particles, is one of the chitosan-induced cellular defense mechanisms that limit virus accumulation in cells.
The results of this work are in line with other available data (Golinowski W, et al., 1981; Roggero P, et al., 1984; Salomon R, 1989; Zhang L, et al., 2001; Trifonova E A, et al., 2007; Reunov A V, et al., 2013) that indicate the possibility of plant virus destruction by the action of hydrolases in vivo.
-
We thank Dr. RV Gnutova and Dr. II Sybiryakova for supplying the antisera used in the work. This work was supported by the Programs for Fundamental Research of the Presidium of the Russian Academy of Sciences "Molecular-cell biology".
-
All the authors declare that they have no competing interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.
-
VP Nagorskaya and LA Lapshina performed the experiments. VN Davydova obtained the chitosan preparation. AV Reunov and IM Yermak conceived and drafted the manuscript. All authors read and approved the final manuscript.

 DownLoad:
DownLoad: