










-
Dear Editor,
The main limitation of the viral pathogenesis study of human gammaherpesviruses Epstein-Barr virus (EBV) and Kaposi sarcoma-associated herpesvirus (KSHV) is the absence of animal model owing to the narrow host tropism of both EBV and KSHV. Murine gammaher pesvirus 68 (MHV68) encodes several genes involved in latency and reactivation that are well conserved among gammaherpesviruses. Additionally the large block of MHV68 genes encoding functions directly involved in virus replication that is also well conserved among all herpesviruses (Barton et al., 2011). The pathogenic similarity of MHV68 infection in laboratory mice and EBV infection in human entitles MHV68 to a valuable small animal model for the study of gammaherpesviral pathogenesis. Similar to that EBV readily immortalizes peripheral blood lymphocytes into lymphoblastoid cell lines (LCLs), MHV68 can also immortalize murine immature B cells into lymphoma B cells (Liang et al., 2011). Our previous work showed that both CD4 and CD8 T cells directly recognize MHV68-immortalized B cells and prevent lymphoma growth in RAG2-/- mice, which is consistent with the data showing that human CD4 or CD8 T cells can prevent the growth of EBV-immortalized LCLs in SCID mice (Lacerda et al., 1996; Liang et al., 2013).
To further define the role of CD4 and CD8 T cells in the control of MHV68-associated lymphoma growth, we depleted CD4 T cells in C57BL/6 mice by injecting anti-CD4 antibody (GK1.5) intraperitoneally at a 2-week interval. Peripheral blood was obtained and subjected to surface staining with CD8a and CD4 antibodies. Flow cytometry analyses showed the efficient depletion of CD4 T cells (Figure 1A). CD4-depleted mice and CD8a-/- mice were intraperitoneally injected with 1×107 MHV68-immortalized SL-1 cells. Wild-type mice and T-cell deficient Nude mice were used as a control. Abdominal tumors appeared 14 to 28 days after injection of cells. Mice were euthanized when tumors reached 3 cm in diameter. As shown in Figure 1B, consistent with the previous observation, MHV68-immortalized SL-1 B cells induced lymphoma in Nude mice, but not in wild-type mice. Surprisingly, no tumor was seen in CD4-depletion mice and CD8a-/- mice. This suggested that mice with either CD4 or CD8 T cell deficiency could still protect against lymphoma growth, which supports our previous observation that either CD4 T cells or CD8 T cells can prevent SL-1-associated tumor growth (Liang et al., 2013).
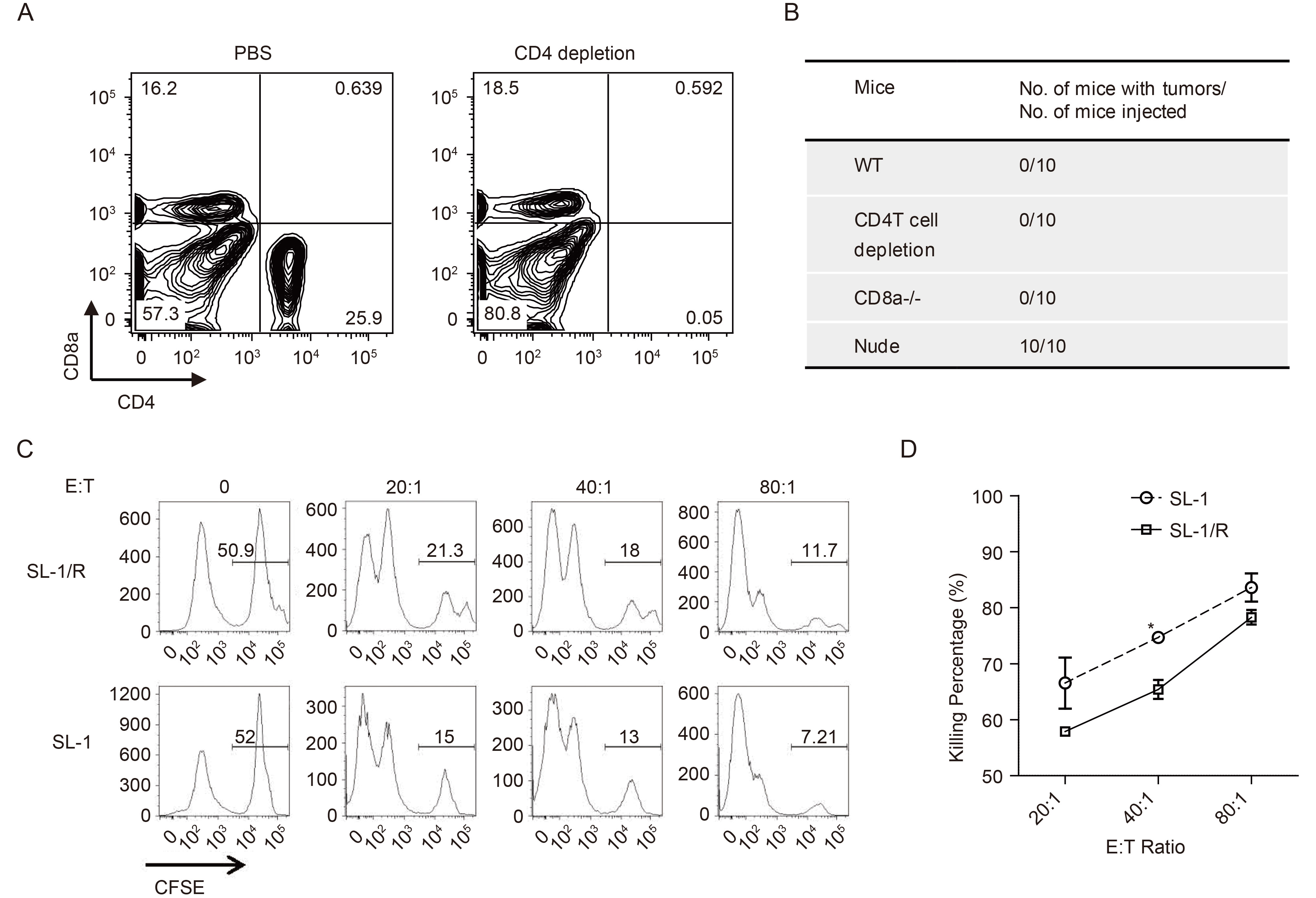
Figure 1. CD95/CD95L involves in the growth control of MHV68-immortalized SL-1 B cells by cytotoxic T cells. (A) Representative flow plot for CD4 T cell depletion. C57BL/6 mice were intraperitoneally injected with 200 μg anti-CD4 antibody (GK1.5) at a 2-week interval. Mice with PBS injection were used as a control. Cells isolated from peripheral blood were subjected to surface staining with CD4 and CD8a antibodies, followed by flow cytometry analyses. (B) Summary of tumor growth in mice injected with MHV68 immortalized SL-1 cells intraperitoneally. (C) Representative histogram plots showing the selective killing of SL-1 and SL-1/R cells by T cells from SL-1-immunized mice. T cells isolated from mice immunized with SL-1 cells were labeled with PKH26 and used as effector cells. Target cells were gated with PKH26 negative and subjected to CFSE staining analyses. CFSEhi (0.5 μmol/L) cells were SL-1 or SL-1/R target cells, where CFSElo (0.01 μmol/L) cells were non-specific splenic cells. The ratio of effector to target cells (E:T) was indicated. (D) Percent specific lysis was calculated by using the formula [1-%CFSEhi (+effectors)/%CFSEhi (no effectors)]×100. *, P < 0.05. Representative data from three experiments are shown. Bars represent standard errors of the means.
Cytotoxic T lymphocytes kill tumor cells by two distinct apoptosis-mediated, fast-acting lytic mechanisms. One is through the granule exocytosis mediated by the secreted lytic proteins perforin and granzyme, the other is involved in the surface membrane CD95 ligand (CD95L) on the cytotoxic T cells, cross-linking with the apoptosis-inducing target cell surface death receptor CD95 (Berke, 1997). CD95/CD95L-mediated apoptosis is a direct major system that both CD8+ cytotoxic lymphocytes and CD4+ cytolytic effector T cells use to eliminate neoplastically transformed cells (Ju et al., 1994; Lowin et al., 1994). The growth of EBV-infected B cells can be regulated by the activated CD4 T cells through apoptosis mediated by CD95/CD95L (Wilson et al., 1998). We recently selected MHV68-immortalized SL lymphoma B cells (SL-1/R) which display CD95 surface downregulation (Tan et al., 2016). We used the parental SL-1 cells with high CD95 surface expression and SL-1/R cells with low CD95 surface expression to examine whether the CD95/CD95L system is involved in SL-1 cell killing by cytotoxic T cells.
We immunized C57BL/6 mice with SL-1 cells (1×107) by intraperitoneal injection, boosted with the second injection after day 15 and isolated pan T cells at day 30 as effector T cells. SL-1 and SL-1/R cells were labeled with 0.5 μmol/L carboxyfluorescein succinimidyl ester (CFSE), respectively. Naïve splenic cells were labeled with a low concentration of 0.01 μmol/L CFSE and used as non-specific target cells. SL-1 and SL-1/R cells were separately mixed with splenic cells at the ratio of 1:1 and used as target cells. The effector T cells isolated from immunized mice were labeled with PKH26 and incubated with target cells for 16 hr. The ratio of effector cells to target cells was 0, 20:1, 40:1 and 80:1. PKH26 negative target cells were further analyzed for the proportion of cells labeled with high CFSE and low CFSE by flow cytometry analyses. Without cytotoxic effector T cells, both nonspecific splenic B cells (low CFSE) and specific target SL-1 (or SL-1/R) cells (high CFSE) were maintained about 50%, at the ratio of 1:1 (Figure 1C). However, consistent with our previous result, pan T cells isolated from immunized C57BL/6 mice display specific cytotoxic killing activity to SL-1 cells, the cell number of SL-1 cells were correspondingly decreased with the increasing ratio of effector to target cells, whereas splenic B cell number was not affected (Figure 1C). Notably, the effector T cells also elicited cytotoxic killing activity to SL-1/R cells, but at a much lower killing rate (Figure 1C, 1D), suggesting that SL-1/R cells with low CD95 surface expression are more resistant to cytotoxic T cell killing, T cell surface membrane CD95L-mediated apoptosis plays a direct role in cytotoxic killing of MHV68-immortalized SL-1 cells. Furthermore, both CD4 T and CD8 T cell isolated from immunized mice expressed similar level of CD95L (CD178). We also performed a similar cytotoxic T cell killing experiment with either CD4 T cells or CD8 T cells and obtained the similar result to that with pan T cells.
In conclusion, our current data demonstrate that MHV68-immortalized SL-1 cells can be recognized and controlled by specific cytotoxic T cells through CD95/CD95L-mediated apoptosis. This is in agreement with that CD4 T cells control the growth of EBV-infected cells through CD95/CD95L-mediated apoptosis, which suggests that the growth control of gammaherpesvirus-associated lymphoma cells by cytotoxic T cell shares conserved mechanism.
HTML
-
This work has been financially funded by grants from the Natural Science Foundation of China (81371825), the Ministry of Science and Technology of China (2016YFA0502100), and the Chinese Academy of Sciences “100 talents” program (2060299). The authors declare that they have no conflict of interest. All animal experiments were performed in strict accordance with the regulations in the Guide for the Care and Use of Laboratory Animals issued by the Ministry of Science and Technology of the People's Republic of China. All efforts were made to minimize pain and suffering. The protocol was approved by the Institutional Animal Care and Use Committee of the Institut Pasteur of Shanghai.

 DownLoad:
DownLoad: