









-
The host immune response system is thought to be involved in the control of HIV-1 infection. During acute HIV-1 infection, the viral load rises to a peak, which leads to activation of the cellular immune system (Kahn and Walker, 1998). This high-level plasma viremia typically declines dramatically once HIV-1-specific cytotoxic T lymphocyte (CTL)-mediated immune responses are mounted, probably reflecting the strong antiviral activities of these cells during acute infection (Borrow et al., 1994; Koup et al., 1994). The dramatically accelerated progression of simian immunodeficiency virus disease that is noted to occur in rhesus macaques following the depletion of CTLs also provides evidence for the importance of CTLs in the control of viruses (Schmitz et al., 1999). Moreover, enhanced ability of HIV-1-specific CTLs to inhibit virus replication in autologous CD4+ T cells was demonstrated in a rare subpopulation of HIV-1-infected individuals (Saez-Cirion et al., 2007; Chen et al., 2012), although these cells actually had low res-ponses as measured at baseline by IFN-γ enzyme-linked immunospot (ELISpot) assays (Pereyra et al., 2008). These individuals maintain levels of viremia below the limit of detection by current assays for long periods in the absence of therapy (Cao et al., 1995; Deeks and Walker, 2007) and are referred to here as elite controllers. Similarly, HIV-1-specific neutralizing antibodies (NAbs) are often undetectable or at low titer during acute infection (Moore et al., 1994; Moog et al., 1997) and in elite controllers (Harrer et al., 1996; Bailey et al., 2006; Pereyra et al., 2008), suggesting that antibodies do not play a major role in viral control. These findings constitute a striking contrast to progressive HIV-1 infection, where high levels of virus replication occur in spite of the presence of strong HIV-1-specific CTL and NAb responses. In these progressive patients, HIV-1-specific CTL responses are of higher magnitude and more broadly directed, as measured by the assessment of IFN-γ production (Pereyra et al., 2008), and HIV-1-specific NAbs are at higher titer, as measured by in vitro neutralization of cell-free viruses (Bailey et al., 2006; Pereyra et al., 2008). The ability of these immune responses to inhibit virus replication in autologous CD4+ T cells, the ultimate function likely required of protective responses, has rarely been measured.
It has been recently reported that antibody-dependent cell-mediated virus inhibition (ADCVI), a measure of the combined potential of antibody and effector cells dependent on an Fc-Fc receptor (FcR) interaction, was able to contain virus infection rather than cause target cell death (Forthal et al., 2006). Passive infusion of neutralizing monoclonal antibodies has been demonstrated to prevent virus infection in rhesus macaques (Emini et al., 1992; Mascola et al., 2000; Hessell et al., 2007). Furthermore, antibody responses to vaccines have correlated with improved outcomes in macaques after virus challenge (Schmitz et al., 2005; Someya et al., 2005).
Given that a cellular immune response is well documented to contribute to control of HIV-1 infection, it is possible that enhanced antiviral efficacy against HIV-1 could be gained by a combination of NAbs and cell-mediated immunity (Hessell et al., 2007; Walker and Burton, 2008). In this study, we examined the potency of CTLs and natural killer cells (NKs) from HIV-1-infected individuals, in combination with engineered NAbs, to inhibit infectious virus replication in autologous CD4+ T cells. Our data indicated that both CTLs and antibody-dependent NK-mediated immunity contribute to viral containment.
-
HIV-1-infected individuals were recruited on the basis of viral load from outpatient clinics at local hospitals, following institutional review board approval and written informed consent. Elite controllers were defined as having HIV-1 RNA below the level of detection for the respective available ultrasensitive assay (e.g., < 75 RNA copies/mL by bDNA or < 50 copies by ultrasenstive PCR) without antiretroviral therapy. Untreated chronic progressors were recruited locally on the basis of viral load above 10, 000 RNA copies/mL plasma.
-
The primary CXCR4-utilizing HIV-1 strain X4 92HT599 and CCR5-utilizing HIV-1 strain R5 91US056 were obtained from the AIDS Research and Reagent Program, Division of AIDS, NIAID, NIH (Bethesda, MD, USA). The titers of the virus stocks were 2.57 × 106 TCID50/mL and 2.14 × 106 TCID50/mL, respectively.
-
IgG1 b12 is an engineered monoclonal human antibody that recognizes an epitope overlapping the CD4-binding site of gp120 (Burton et al., 1994; Binley et al., 2004; Nie et al., 2014). LALA, a variant of b12, was created by site-directed mutagenesis (Hezareh et al., 2001). LALA retains antigen binding but binds weakly to human Fcγ receptors, triggers antibody-dependent cellular cytotoxicity (ADCC) with lower efficiency, and binds and activates human complement weakly (Hessell et al., 2007). IgG1 DEN3 is an isotype control monoclonal antibody specific for Dengue virus.
-
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood of HIV-1-infected individuals by Ficoll-Hypaque density gradient centrifugation. Bulk CD8+ T cells and NKs were isolated from PBMCs by negative selection (Dynabeads, Thermo Fisher Scientific, Waltham, Massachusetts, USA). The isolated CD8+ T cells and NKs were of up to 98% purity as validated by flow cytometry. The generated CD8+ T cells responding to viral proteins were fine-mapped using optimal peptide pools in IFN-γ ELISpot assays.
-
IFN-γ ELISpot assays were performed as described by Addo et al. (2003) , using optimally defined epitope peptide pools. To calculate the number of specific spot-forming cells (SFC), the number of spots in the negative control wells was subtracted from the number of spots counted in each well. Negative controls were always 30 SFC per 106 input cells or fewer.
-
Virus neutralization was performed using the method described by Trkola et al. (1995) with slight modifications. Briefly, CD4+ T cells were generated from freshly isolated PBMCs by the addition of CD3: CD8 bispecific monoclonal antibody (Wilson et al., 1995; Wong et al., 1997). After 5–7 days, greater than 98% of these cells coexpressed CD3 and CD4 by flow cytometric analysis (data not shown). Antibodies were adjusted to 200 μg/mL, and eight twofold serial dilutions were prepared. An aliquot (50 μL) of each dilution step was transferred to four replicate wells of a 96-well culture plate. The virus inoculum was adjusted to 2000 TCID50/mL, and a 50-μL (100 TCID50) aliquot was incubated with antibodies (50 μL) for 1 h at 37 °C. The calculated neutralization titers refer to the antibody concentrations present during this preincubation step. The antibody-virus mixture was then diluted twofold by addition of 100 μL of CD4+ T cells at a density of 1 × 106/mL, and the cultures were incubated for 7 days. The cultures were collected and treated with 0.5% Triton, and the p24 antigen concentration was determined by a standard quantitative enzyme-linked immunosorbent assay (HIV-1 p24 ELISA kit, PerkinElmer, Inc., Waltham, Massachusetts, USA). As the antibody-virus inoculum was not washed away from the cells at any stage, control wells containing 50 μL of virus and 150 μL of medium were included to provide an estimate of the amount of input p24 antigen that carried over into the p24 antigen assay for virus production. This value was subtracted from all the test results. The antibody concentrations (μg/mL) causing 50%, 90%, and 99% reduction in p24 antigen production were determined by linear regression analysis; these are the ID50, ID90, and ID99 values.
-
Inhibition of viral replication was assessed in a previously established assay system (Yang et al., 2002; Forthal et al., 2006; Hessell et al., 2007) with slight modifications. Briefly, autologous CD4+ T cells were infected with designated HIV-1 strains at a multiplicity of infection (MOI) 0.01 for 4 h at 37 °C, washed twice, and plated at 5 × 104 cells per well in a 96-well round-bottom plate. Various dilutions of b12, LALA mutant, or isotype control DEN3 antibodies were added to virally infected cells alone or along with effector cells, either CD8+ T cells or NKs, at a ratio of 1:1 or as otherwise specified in a total of 200 μL RPMI 1640 medium containing 50 U/mL interleukin-2 (IL-2). After 7 days of incubation at 37 °C in 5% CO2, supernatant was collected and p24 antigen was quantified by a standard quantitative enzyme-linked immunosorbent assay (HIV-1 p24 ELISA kit, Dupont, Boston, MA, USA). Virus inhibition was calculated as follows: Percentage inhibition = 100 × (1 - p24p/p24n), where p24p and p24n are the concentrations of p24 in supernatant from wells with or without a source of effectors, respectively.
-
Mann-Whitney tests were performed using GraphPad Prism version 4.0a, and P-values of P < 0.05 were considered significant.
-
Using two primary HIV-1 isolates, X4 92HT599 and R5 91US056, we first tested the ability of monoclonal antibody b12 and its mutated Fc- and compliment-binding variant LALA to neutralize viruses using CD4+ T cells from elite controllers as indicator cells. The outgrowth of autologous viruses in CD4+ T cells from the elite controllers was markedly delayed, and there were no autologous viruses grown out from the tested subjects during the period of the assay. Representative neutralization titration curves are depicted in Figure 1A.
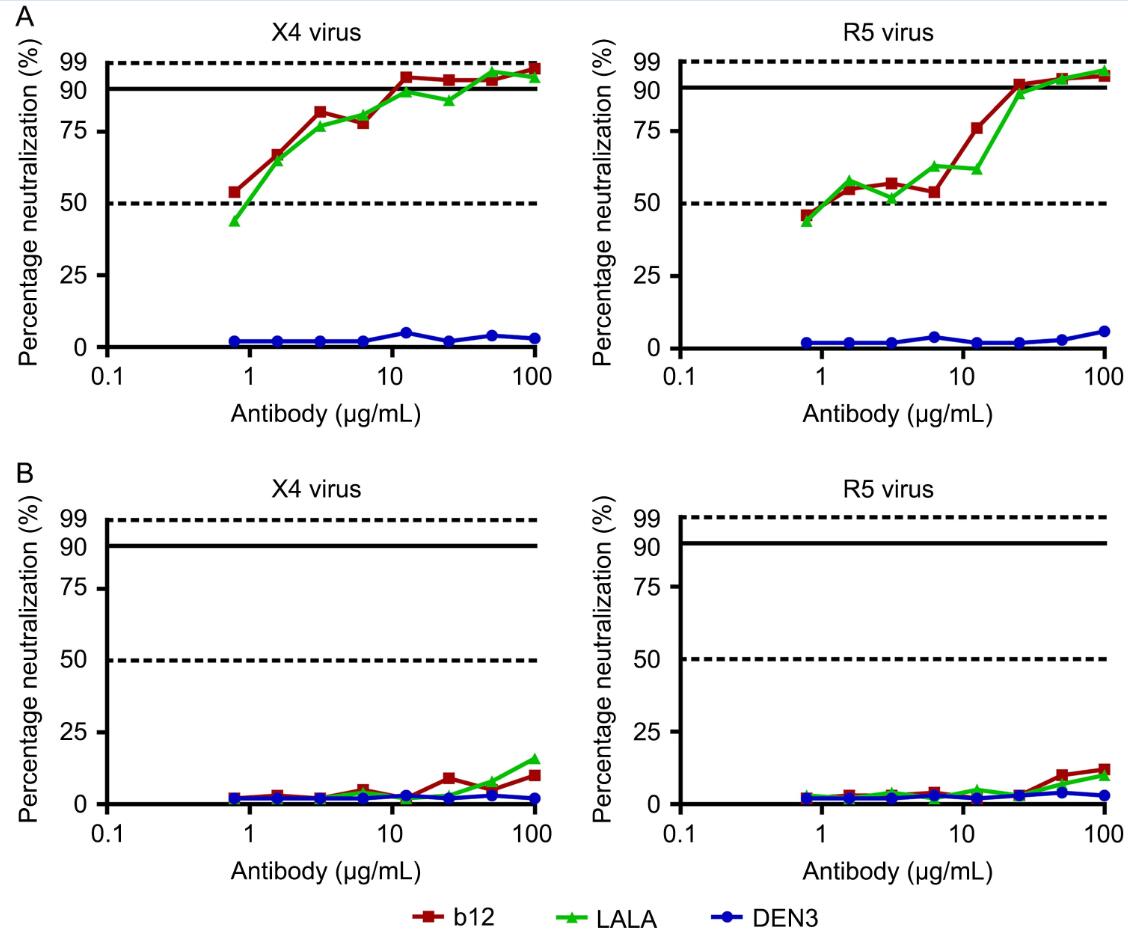
Figure 1. Neutralization of HIV-1 by antibodies. (A) Monoclonal antibodies were serially diluted and incubated with 100 TCID50/mL of the indicated primary viruses for 1 h at 37°C prior to adding to CD4+ T cells. (B) Monoclonal antibodies were serially diluted and incubated with CD4+ T cells that had been infected with HIV-1 at an MOl of 0.01 4 h earlier. The extent of virus replication was monitored by measurement of p24 antigen production after 7 days. The percentage of neutralization is plotted against antibody concentrations. ID50, ID90, and ID99 values are indicated
-
The ID90 for both X4 and R5 virus for b12 were in the range of 10 to 25 μg/mL, and LALA was comparable to b12 with an ID90 in the range of 10 to 30 μg/mL. These data showed that both b12 and the LALA variant demonstrated neutralization of cell-free X4 and R5 primary isolates, but for this inoculum of virus a relatively high concentration of antibody was required to achieve 95% neutralization. Moreover, the data showed that both b12 and LALA, which lacks FcγR and complement binding functions compared with b12, mediate comparable neutralization of these primary viruses.
Next, we tested the ability of these antibodies to contain viruses in virally infected CD4+ T cells from the elite controllers in whom outgrowth of autologous viruses in CD4+ T cells was markedly delayed. CD4+ T cells were infected with primary HIV-1 isolates prior to the addition of antibodies. Designated concentrations of b12, LALA mutant, or isotype control DEN3 antibodies were added to virally infected cells. The p24 antigen production in supernatant was evaluated and expressed as the percentage of neutralization (Figure 1B). Both b12 and LALA failed to effectively control viral replication in infected CD4+ T cells as the p24 antigen production in supernatant steadily increased during the period of 7 days even with antibody at 100 μg/mL, far in excess of the range estimates for concentration of an antibody produced in vivo. These data suggest that although this antibody is potent in neutralizing free virus, its ability to neutralize virus spread once infection has occurred is very limited, which is likely due to cell-cell spread of virus in this in vitro culture system, but may also be related to production of high numbers of progeny virions that overwhelm the available antibody binding sites.
-
We next tested the ability of bulk CD8+ T cells from HIV-1 elite controllers and chronic progressors to inhibit virus replication in the same cell culture system, using autologous CD4+ T cells preincubated for 4 h with the same HIV-1 strains at the same MOI (0.01) at which no effect was observed using the antibodies alone (Chen et al., 2009). For these experiments, we chose elite controllers and chronic progressors in whom outgrowth of autologous viruses in CD4+ T cells was markedly delayed, allowing us to use a fixed inoculum of primary viruses in these experiments. As shown as an example in Figure 2A, addition of autologous bulk CD8+ T cells from an elite controller to virally infected CD4+ T cells at a CD8+ to CD4+ T cell ratio of 1 to 1 resulted in marked inhibition of virus replication over 7 days in culture, whereas weak inhibition of virus replication was observed in coculture of virally infected CD4+ T cells with autologous bulk CD8+ T cells from a progressor (Figure 2B).
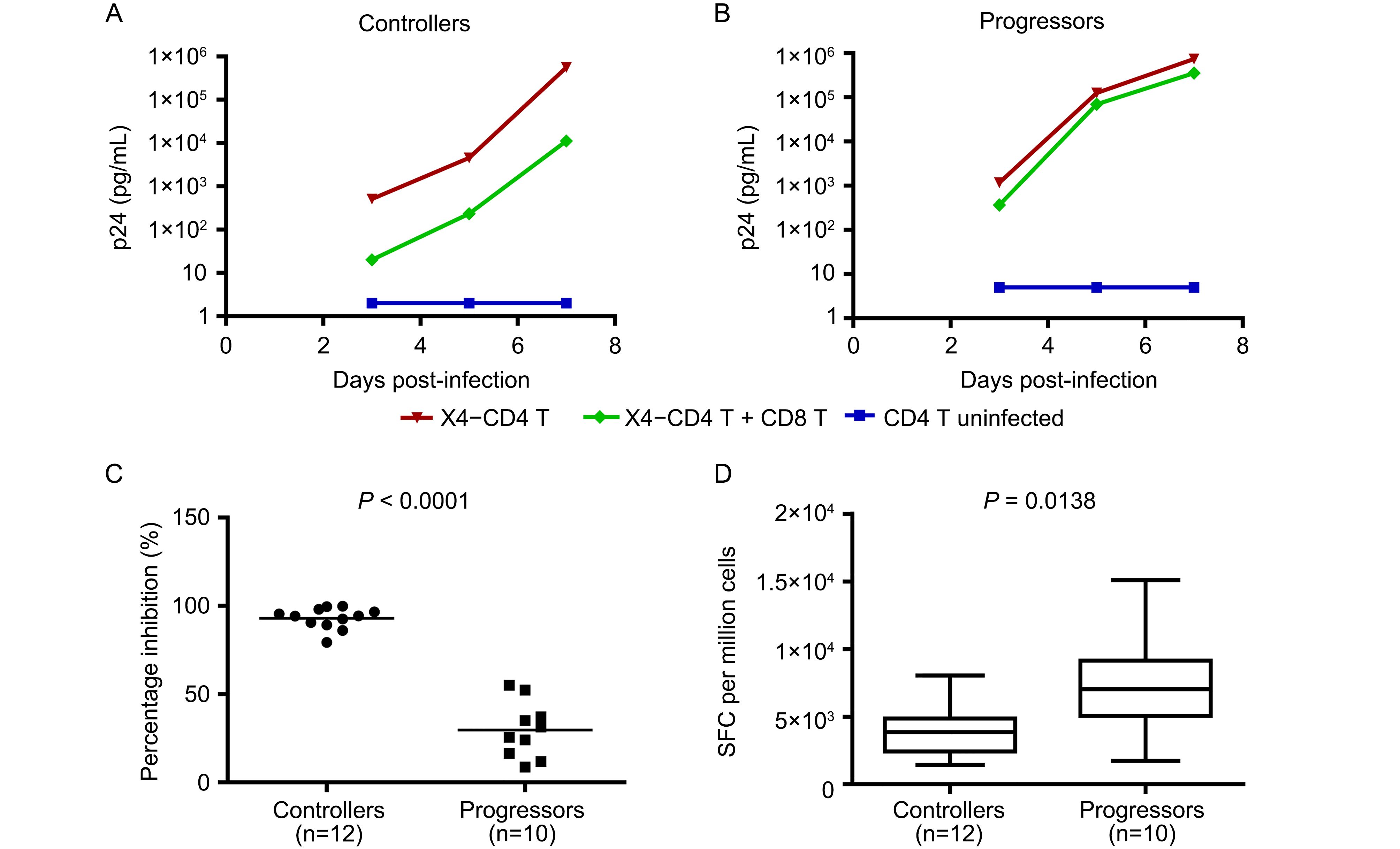
Figure 2. Suppression of HIV-1 by CTLs. The antiviral ability of bulk CD8+ T cells was assessed by an in vitro virus inhibition assay, by adding CTLs to autologous CD4+ cells infected with X4 virus at an MOI of 0.01 4 h earlier. (A) Bulk CD8+ T cells from an elite controller effectively inhibited HIV-1 replication at a 1:1 ratio of CD8+ to CD4+ T cells during a period of 7 days. Results from one experiment are shown as an example. (B) Bulk CD8+ T cells from a progressor had weak antiviral efficacy at a 1:1 ratio of CD8+ to CD4+ T cells. Results from one experiment are shown as an example. The control uninfected CD4+ T cells showed that there were no autologous viruses grown out from the tested subjects during the period of the assay. (C) Bulk CD8+ T cells from the controllers (n = 12) were more effective than those from the progressors (n = 10) in control of virus replication. Data are expressed as the percentage of inhibition, calculated as 100×(1 - p24 with effectors/p24 without effectors) at day 7 in culture. (D) IFN-γ ELISpot study showed HIV-1-specific CTL responses in bulk CD8+ T cells from elite controllers (n = 12) were weaker than those from chronic progressors (n = 10). In the box and whisker plot, the line indicates the mean, the box indicates lower quartile and upper quartile, the whisker indicates the minimal and the maximal numbers, Statistical comparisons were made using the Mann-Whitney test.
We next expanded these studies to include 12 elite controllers and 10 chronic progressors, examining the ability of their CTLs to inhibit the same strain of HIV-1 in autologous CD4+ T cells. Marked inhibition of virus replication was observed at a CD8+ T to CD4+ T cell ratio of 1 to 1 for the elite controllers whereas there was weak inhibition of virus replication by bulk CTLs from the chronic progressors (Figure 2C). The difference was highly statistically significant (P < 0.0001). The magnitude of HIV-1-specific CTL responses in bulk CTLs was evaluated by optimally defined epitope pools in IFN-γ ELISpot assays ( Figure 2D). Although the ability of CTLs to retard virus replication was greater in the elite controllers, the magnitude of HIV-1-specific CTL responses as measured in the elite controllers was actually weaker than those in the progressors (3973 ± 1935 vs. 7324 ± 3691 SFC per million bulk CTLs, P = 0.0138). These data indicate that HIV-1-specific CTLs from elite controllers have an enhanced antiviral functional capacity, contributing to the control of virus infection, which is not measured by standard IFN-γ ELISpot assays, suggesting IFN-γ ELISpot approaches may be of limited value in defining CTL functions such as the ability to neutralize virus replication. These data also demonstrate that CTLs from elite controllers can significantly inhibit replication of reference strains of primary isolates in cell culture, whereas a potent NAb, although highly effective when exposed to virus pre-innoculation, is ineffective once initial infection of the cells has occurred.
We next addressed the possibility that enhanced antiviral efficacy could be gained by a combination of CTLs and NAbs. In the infected cell cultures, serially diluted concentrations of NAb b12 were added along with CTLs. However, there was no significantly enhanced neutralization observed with added antibodies even at the highest concentration of 100 μg/mL for CTLs from HIV-1 elite controllers (Figure 3A) or chronic progressors (Figure 3B). Given ADCVI is reported to be dependent on Fc-FcR interaction (Forthal et al., 2006; Hessell et al., 2007), we next determined FcR expression at the surface of the HIV-1-specific CTLs by flow cytometry. Little or no expression of various kinds of FcR was detected on the surface of the HIV-1-specific CTLs (data not shown). Using a previously reported assay (Liu et al., 2009), degranulation of intracellular vesicles (CD107a expression) mediated by Fc-FcR interaction was examined. There was no CD107a upregulation on the CTLs after stimulation with IgG-loaded p815 cells compared with stimulation with p815 cells alone or medium control (data not shown). These data may explain the above finding that enhanced antiviral efficacy against HIV-1 could not be gained by ADCVI mediated by HIV-1-specific CTLs.

Figure 3. Inhibition of HIV-1 by CTLs in the presence of NAbs. Autologous CD4+ cells infected with X4 virus at an MOI of 0.01 for 4 h. NAb b12 at 100 μg/mL was added along with bulk CD8+ T cells from (A) elite controllers (EC, n = 12) or (B) chronic progressors (CP, n = 10) at a 1:1 ratio of CD8+ to CD4+ T cells during a period of 7 days of culture. Data are expressed as the percentage of inhibition. There was no significantly enhanced inhibition to the bulk CD8+ T cells with added NAb b12.
-
NKs have been reported to serve as main effector cells and in combination with NAbs to mediate ADCVI as described previously (Forthal et al., 2006; Hessell et al., 2007; Liu et al., 2009; Asmal et al., 2011). We next evaluated ADCVI of NKs from virally infected individuals in combination with NAbs. Autologous CD4+ T cells were infected with primary X4 virus and cocultured with NKs alone or along with designated concentrations of antibodies. As shown as an example in Figure 4A, addition of autologous NKs to virally infected CD4+ T cells resulted in significantly enhanced inhibition of virus replication over 7 days in culture containing 50 U/mL IL-2 and this enhanced inhibition was mediated by antibody b12 but not by LALA or the isotype control monoclonal antibody DEN3. From the study of the 12 elite controllers and 10 chronic progressors, weak inhibition of primary X4 virus was observed by NKs alone from the controllers (Figure 4B) or progressors (Figure 4C), which may be mediated by direct killing of NKs through activation of receptors on NKs that interact with major histocompatibility complex class I molecules on HIV-1-infected CD4+ T cells (Bonaparte and Barker, 2004; Vieillard et al., 2005). However, significantly enhanced inhibition (P < 0.01) was gained by addition of antibody b12 at a concentration of 100 μg/mL to NKs for both the controllers ( Figure 4B) and progressors (Figure 4C) whereas there was no statistical difference in the antiviral activity of NKs alone (P = 0.5704) or antibody-mediated enhanced inhibition (P = 0.8336) bewteen the controllers and progressors. Moreover, this impact by ADCVI was mediated by antibody b12 but not by LALA, further demonstrating that antiviral potency by ADCVI is dependent on Fc-FcR interaction.
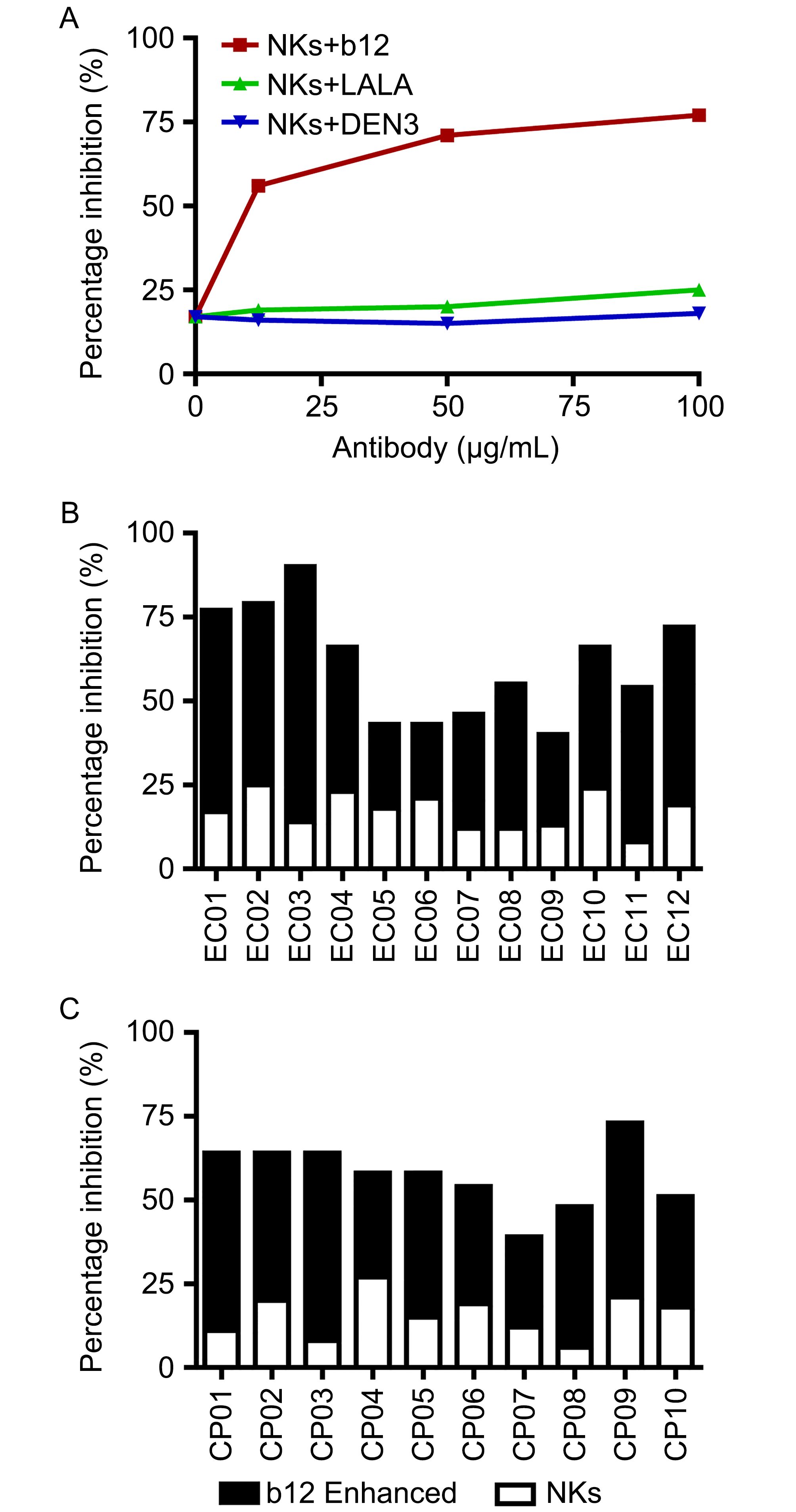
Figure 4. Inhibition of HIV-1 by ADCVI. Autologous CD4+ T cells were infected with primary X4 virus at an MOI of 0.01 for 4 h and then incubated with NKs alone or along with designated concentrations of b12, LALA mutant, or isotype control DEN3 antibodies. The extent of virus replication was monitored by measurement of p24 antigen production after 7 days. (A) The percentage of inhibition of an example experiment was plotted against antibody concentration. (B) ADCVI was mediated by NKs from elite controllers (EC, n = 12) in the presence of NAb b12 at 100 μg/mL. (C) ADCVI was mediated by NKs from chronic progressors (CP, n = 10) in the presence of NAb b12 at 100 μg/mL.
-
The relative contributions of various immune mechanisms to control of ongoing viral replication remain elusive. These data, employing a functional assay that measures the ability of effector cells, neutralizing monoclonal antibodies, and the combined power of these effectors to inhibit viral replication of primary HIV-1 isolates in autologous CD4+ T cells, indicate that NAb b12 is highly potent in neutralizing free viruses but is limited in containment of cell-cell spread of viruses once infection has occurred. In contrast, bulk CTLs from HIV-1 controllers are effective in containment of viruses under conditions where little or no effect is seen from NAbs. Moreover, bulk CTLs from HIV-1 controllers are more effective in suppression of virus replication than those from progressors. However, the same b12 NAb is able to contain viruses by ADCVI. Importantly, these data demonstrate that ADCVI is mediated by NKs and dependent on an Fc-FcR interaction, suggesting innate immune mechanisms such as NKs or monocytes/macrophages that generally express FcRs may contribute to control of ongoing viral replication by ADCVI in the presence of functional antibodies. However, these data and previous findings that the high-level plasma viremia declines dramatically once HIV-1-specific CTL responses are mounted (Borrow et al., 1994; Koup et al., 1994) probably reflect that the HIV-1-specific CTLs are the major effector cells in containment of viruses whereas the mounted NK response during acute HIV-1 infection is not enough to contain viruses.
The goal of HIV-1 vaccination strategies is to have an immunologic barrier in place that will prevent infection or limit virus replication once infection occurs, thereby limiting symptoms of disease caused by virus infection (Wang et al., 2016). A priority in HIV-1 vaccine development has been the generation of NAbs, ideally to provide sterilizing immunity, but this has remained an elusive goal. Only one antibody-based AIDS vaccine has been taken through efficacy trials thus far, using the gp120 subunit protein as an immunogen, and this vaccine candidate did not elicit antibodies neutralizing field isolates of HIV-1, did not prevent infection, and did not affect subsequent viral load (Flynn, 2005). Although the recent RV144 Thai trial showed moderate protection against viral acquisition (Gilbert et al., 2011), the protection mechanisms remain unclear (Kim et al., 2015). The most important second line of defense is the cellular immune response, including NKs and CTLs as the presumed major effector arms. Together with previous findings (Yang et al., 1996; Yang et al., 1997; Yang et al., 2003; Chen et al., 2009), our studies have demonstrated that both CTLs alone and NKs in the presence of functional antibodies can suppress HIV-1 replication in vitro. However, not all CTL responses created in natural HIV-1 infection are equally potent in terms of the ability to inhibit virus replication. The limitation of this study is that all these antiviral factors were assessed in vitro. It is critical to define in vivo HIV-1-specific CTL responses for viral control during natural infection and vaccine protection against HIV-1.
There is a remarkable subset of HIV-1-infected individuals, here named elite controllers, who are able to maintain a state of apparent durable control of HIV-1 replication without the need for antiviral therapy, providing clear evidence that durable containment of HIV-1 in the absence of therapy is possible. Some evidence for the involvement of CTL responses (Migueles et al., 2002; Zimmerli et al., 2005; Betts et al., 2006) was documented for the mechanisms of virus control in these cases. In this study, a virus suppression assay was used to assess the antiviral potential of CTL responses from these subjects versus chronic progressors. As compared with chronic progressors, bulk CTLs from elite controllers had higher capacities to inhibit virus replication in infected autologous CD4+ T cells, which is consistent with the finding by Saez-Cirion and his colleagues that HIV-1 controllers rather than progressors exhibited potent CTL capacity to suppress HIV-1 infection ex vivo (Saez-Cirion et al., 2007). Factors that potentially modulate protective CTL responses include, among others, polyfunctionality, antigen sensitivity or functional avidity, proliferative capacity, loading of lytic granules, ex vivo expression of perforin, T cell antigen receptor (TCR) clonotypes, and properties of the interactions among TCRs, viral peptide, and the major histocompatability complex (Chen et al., 2012). Collectively, this evidence strongly argues that potent CTL responses directed at HIV-1 are causally related to HIV-1 control in at least some controllers. However, many of these studies are by necessity correlative in nature, making it difficult to rule out the possibility that the preservation of potent HIV-1-specific CTL responses is a consequence rather than a cause of HIV-1 control. Although NKs might exert innate antiviral immunity in acute HIV-1 infection, the mounted NK cell response during acute HIV-1 infection does not appear to be enough to contain viruses. The role of NKs in chronic HIV-1 infection in elite controllers and progressors remains unclear. Unraveling the mechanisms of HIV-1 control in these cases should be a high priority because this may facilitate the design of immunogens able to elicit the corresponding protective responses.
It has been reported that certain engineered monoclonal antibodies and primary antibodies from HIV-1-infected individuals, in the presence of effector cells, were able to inhibit HIV-1 replication in vitro (Forthal et al., 2001; Forthal et al., 2006) and modulate viremia in vivo dependent on Fc-FcR interactions (Hessell et al., 2007). This control of virus replication involving not only cytolytic but also noncytolytic mechanisms was distinguished from ADCC and termed ADCVI by Forthal and his colleagues (Forthal et al., 2006). Given that HIV-1-specific CTLs tested in this study do not express FcRs (data not shown), they would be unlikely to mediate ADCVI, although they showed highly potent ability to inhibit virus replication and limit cell-cell spread of viruses. NKs were reported to serve as the effector cells in ADCVI (Forthal et al., 2001; Forthal et al., 2005; Forthal et al., 2006) because NKs express various kinds of FcRs and can therefore interact with antibodies to modulate antiviral activity. It is to be expected that there are other subsets of cells, such as monocytes/macrophages, which express FcRs, involved in ADCVI and that contribute to viral control. ADCVI also allows the possibilty that antibodies, passively transferred via a viral vector, could be competent to control viruses in the presence of effector cells, overcoming the barrier that antibodies that neutralize HIV-1 are often absent or at low titer in natural HIV-1 infection. The protection mechanisms for HIV-1 containment need to be further investigated with broader CTLs, NAbs, NKs, or other factors in the context of acute infection.
Taken together, our study, for the first time to our knowledge, provides a systemic comparison of three major players, NAbs, CTLs, and NKs, in an autologous system to assess neutralization, cytotoxicity, and ADCVI on HIV-1 replication in CD4+ T cells, and shows that no single arm thus far can be defined as the correlate of complete protection of HIV-1 infection. Alternative factors or combinations of multiple factors must be pursued. Our findings provide an important consideration for HIV-1 control and vaccine success that may be crucially dependent on the combination of antibody and cell-mediated immunity, as well as the innate and adaptive immunity.
-
This work was supported by the Fund for Jiangsu Specially-Appointed Professor (2014JSTPJS-53), the Innovation Fund from National Center for AIDS/STD Control and Prevention, China CDC, Capital Medical University Key Laboratory Project (2-03-02-BJYAH2016006), and China’s 12th Five-Year Major Project on the Prevention and Treatment of AIDS. Dr. Dennis Burton (the Scripps Research Institute) kindly provided b12, b12 LALA, and Den3 monoclonal antibodies. The HIV-1 strains X4 92HT599 and R5 91US056 were obtained from the AIDS Research and Reagent Program, Division of AIDS, NIAID, NIH.
-
The authors declare that they have no conflict of interest. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee.
-
HBC and CJ designed and supervised the experiments, TT, JBZ, TL, DM, WL, BS, and TZ collected samples and carried out the experiments, HBC, CJ, TT, and JBZ analyzed the data and wrote the paper. NL and HW provided helpful suggestions about the study. All authors read and approved the final manuscript.
In vitro inhibition of HIV-1 replication in autologous CD4+ T cells indicates viral containment by multifactorial mechanisms
- Ting Tu 1,2,# ,
- Jianbo Zhan 3,# ,
- Danlei Mou 4 ,
- Wei Li 4 ,
- Bin Su 4 ,
- Tong Zhang 4 ,
- Tao Li 1 ,
- Ning Li 4 ,
- Hao Wu 4 ,
-
Cong Jin
2,,
 ,
, -
Huabiao Chen
1,,
- Received Date: 05 April 2017
- Accepted Date: 27 July 2017
- Published Date: 15 September 2017
Abstract: HIV-1-specific cytotoxic T lymphocytes (CTLs) and neutralizing antibodies (NAbs) are present during chronic infection, but the relative contributions of these effector mechanisms to viral containment remain unclear. Here, using an in vitro model involving autologous CD4+ T cells, primary HIV-1 isolates, HIV-1-specific CTLs, and neutralizing monoclonal antibodies, we show that b12, a potent and broadly neutralizing monoclonal antibody to HIV-1, was able to block viral infection when preincubated with virus prior to infection, but was much less effective than CTLs at limiting virus replication when added to infected cell cultures. However, the same neutralizing antibody was able to contain viruses by antibody-dependent cell-mediated virus inhibition in vitro, which was mediated by natural killer cells (NKs) and dependent on an Fc-Fc receptor interaction. Meanwhile, bulk CTLs from HIV-1 controllers were more effective in suppression of virus replication than those from progressors. These findings indicate that control of HIV-1 replication in activated CD4+ T cells is ineffectively mediated by neutralizing antibodies alone, but that both CTLs and antibody-dependent NK-mediated immune mechanisms contribute to viral containment. Our study systemically compared three major players in controlling HIV-1 infection, CTLs, NAbs, and NKs, in an autologous system and highlighted the multifactorial mechanisms for viral containment and vaccine success.

 DownLoad:
DownLoad: