









-
Human endogenous retrovirus (HERV) is reported to account for about 8% of the human genome, and its sequence is highly conserved (Jern et al., 2004). HERV-W is a typical representative of the HERV family. A single complete open reading frame (ORF) that putatively encodes the HERV-W envelope (env) is located on human chromosome 7 and can be transcribed and translated into the envelope protein syncytin-1 (Frendo et al., 2003). HERV-W env is not only highly expressed in the placenta but also has fusogenic properties evident in normal trophoblast development in healthy humans (Mallet et al., 2004). However, many more studies have shown that HERV-W env contributes to neuropsychological diseases, including schizophrenia (Karlsson et al., 2004; Huang et al., 2006; Huang et al., 2011), Alzheimer’s disease (AD) (Nakamura et al., 2012), and multiple sclerosis (MS) (Perron et al., 2005; Perron et al., 2012; Perron et al., 1997). Furthermore, these diseases may be associated with brain inflammation (Zipp et al., 2006).
Although the brain is considered an “immunologically privileged site”, mainly because of the blood-brain barrier (BBB), immunological reactions can occur in the CNS. Brain inflammation has been implicated in the pathology of neuropsychological diseases such as stroke, Parkinson’s disease (PD), and human immunodefi- ciency virus-associated dementia (Klegeris et al., 2007; Nakamura et al., 2012; Zipp et al., 2006). In addition, microglia are recognized as immunocytes in the CNS, and their function is to protect the brain from pathogenic bacteria and viruses, and the inflammatory response to these microorganisms (Maeda et al., 2016). A number of studies have shown cellular migration in CNS (Bao et al., 2016). After CNS damage, microglia at the lesion will enter pro-inflammatory (Draheim et al., 1999; Moussaud et al., 2009) and anti-inflammatory (Czeh et al., 2011; Hanisch et al., 2007) sites to promote repair neurocytes proliferation and migration. Microglia, a type of glial cell, produce two subtypes of identified nitric oxide synthase (NOS); one is a constitutive neuronal NOS (nNOS) and the other one is an inducible NOS (iNOS), which promotes the production of additional NO in response to various stimuli (Ghasemi et al., 2014; Li et al., 2009). NO, a free radical molecule, is synthesized from L-arginine by NOS in many mammalian cells and tissues, especially in the brain. At lower concentrations (pmol), nNOS-derived NO exerts beneficial effects on the body by playing vital roles such as antibacterial and antiviral activities (Ghasemi et al., 2014; Kumar et al., 2017). At higher concentrations (μmol), iNOS-derived NO is involved in the pathogenesis of various neuropsychological diseases, including neuroinflammatory diseases, schizophrenia, epilepsy, neurodegenerative diseases, and stroke (Kumar et al., 2017).
Numerous factors have been associated with neuropsychiatric disorders; however, the cause of these diseases remains unclear (Yolken et al., 1995). As discussed above, both HERV-W env and NO contribute to the pathogenesis of neurological disorders such as MS, AS, and schizophrenia (Asiimwe et al., 2016; Ghasemi et al., 2014). Through immunohistochemistry, microglia have been shown to express HERV-W env (Mameli et al., 2007; Antony et al., 2004). However, questions remain: is there an association between HERV-W env and changes in NO levels in microglia in the CNS, and what factors directly lead to changes in NO levels? Answers to these questions have not been reported in the literature.
In this study, by measuring the expression of iNOS and changes in microglia migration, we showed that overexpression of HERV-W env promoted inflammation in microglia. Results of this study can suggest new approaches to therapeutic interventions for neuropsychological diseases.
-
pCMV-env was constructed in a previous study (Huang et al., 2011). Luciferase reporter gene plasmids containing iNOS promoters 8 kb, 7 kb, 6 kb, 4 kb, 3 kb, 2 kb, and 1 kb in length were given as gifts by Professor Ying Zhu, Wuhan University, China (Zou et al., 2007).
-
A common human microglia cell line, CHME-5, given as a gift by Professor Wenzhe Huo (Department of Anatomy and Molecular Biology, Temple University, USA), were cultured in Dulbecco’s modified Eagle’s medium (Gibco BRL) supplemented with 10% fetal bovine serum (Gibco BRL), 2 mmol/L GlutaMAX (Gibco BRL), 10 mmol/L MEM non-essential amino acids (Invitrogen), 100 U/mL penicillin, and 0.1 mg/mL streptomycin at 37 °C in a humidified 5% CO2 incubator (Janabi et al., 1995; Shaikh et al., 2012). Transfection was performed in complete medium without antibiotics when the density of cells was about 70%. The plasmids were transfected into cells using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Cells were collected after transfection for 24 h or 48 h. Cell culture supernatants were collected at 6, 12, 24, and 48 h after transfection and immediately stored at –70 °C until use.
-
NO production was indirectly quantified by measuring nitrite levels using a NO Detection Assay Kit (Beyotime, China). A standard curve of nitrite was established, and nitrite in each sample was assessed by mixing 50 μL of culture supernatant with 50 μL of Griess reagent I/II in a 96-well plate. Plates were gently shaken and held at 20 °C for 10 min, and the absorbances at 540 nm were recorded. Nitrite concentrations in the samples were compared with the nitrite standard curve, with nitrite concentrations ranging from 0 to 100 μmol/L.
-
Total RNA was extracted with TRIzol reagent (Invitrogen) from cells and treated with DNase I (Promega, Madison, WI) according to the manufacturer’s protocol. After RNA extraction, cDNA was obtained using a Clontech SMART PCR cDNA Library Construction Kit (Clontech, Palo Alto, CA). Quantitative real-time PCR was performed using DNA SYBR Green PCR Master Mix (Molecular Probes, Eugene, OR, USA) in an iCycler System (Bio-Rad, Hercules, CA, USA), and results were normalized to β-actin levels. Sequences of primers used in real-time quantitative PCR are listed below: iNOS sense, 5′-CAAAGGCTGTGAGTCCTGCAC-3′; iNOS antisense, 5′-ACTTTGATCAGAAGCTGTCCC-3′ (Antony et al., 2007); HERV-W env sense, 5′-CCATGCCGCTGTATGACCAG-3′; HERV-W env antisense, 5′-GGGTTCCCTTAGAAAGACTCCT-3′; β-actin sense, 5′-CATGTACGTTGCTATCCAGGC-3′; and β-actin antisense, 5′-CTCCTTAATGTCACGCACGAT-3′ (Yu et al., 2014).
-
Cells were washed twice with PBS on ice and lysed by adding lysis buffer (Pierce Chemical, Rockford, IL, USA). Supernatants were collected after centrifugation at 14,000 g for 10 min at 4 °C. Protein concentrations were determined using a BCA Protein Assay Kit. For each sample, 40 μg of total protein was separated by 8% SDS-PAGE. Western blotting was performed using rabbit anti-human iNOS antibody (ab3523, Abcam) at a dilution of 1:200. Polyclonal goat anti-rabbit IgG horseradish peroxidase (HRP)-conjugated antibody (cw0103s, CWBiotech) was used as the secondary antibody at a dilution of 1:5000. β-actin (SAB2701711, Sigma-Aldrich), at a concentration of 1:30,000, was used as a control.
-
iNOS activity was detected using a Nitric Oxide Synthase Assay Kit (Beyotime, China) according to the manufacturer’s instructions. S-methylisothiourea sulfate (SMT) (Beyotime, China), a highly selective inhibitor of iNOS, was dissolved in NO synthase detection buffer (1×) for a final concentration of 100 mmol/L. After 24 h of transfection, CHME-5 cells were gently digested with trypsin (0.25% EDTA-trypsin: 1× D-hanks solution = 1: 4) for 2 min and washed with ice-cold PBS three times. The fluorescence intensity of DAF-FM DA, a fluorescent probe for NO, was measured by flow cytometry (FACS Aria III, BD) at excitation and emission wavelengths of 495 nm and 515 nm, respectively. Results are expressed in relative fluorescence activity units (RFUs).
-
Luciferase activities were measured using a Dual Luciferase Reporter Gene Assay Kit (Beyotime, China) according to the manufacturer’s instructions. Twenty-four hours after co-transfection with luciferase reporter plasmids and pCMV or pCMV-env, CHME-5 cells were washed two times using 1× PBS and lysed by cell lysis buffer for 10 min. Firefly luciferase and Renilla luciferase activities were detected, and the latter, an internal control, was used to eliminate the impact of cell number and transfection efficiency. Results are expressed in relative luciferase activity units (RLUs).
-
Migration of the transfected cells was measured by Transwell assay according to the manufacturer’s protocol. For the cell migration assay, cells were diluted to 1.0 × 105/mL and cultured in 100 μL of cell suspension without fetal bovine serum (FBS) in the upper chamber, and 600 μL of culture medium with 10% FBS was used to create an osmotic pressure difference to promote cell migration to the lower chamber of the Transwell insert (Corning). After 12 h of culture in a 37 °C incubator, the cells that did not migrate were removed with a cotton swab. Migrating cells were fixed with 90% ethanol for 15 min and then stained with crystal violet for 20 min. Then, the migrated cells in at least four fields were observed and counted under a high magnification (400×) inverted microscope. A wound healing assay was also performed. Procedures for the assay are described elsewhere in detail (Zhou et al., 2017; Zhang et al., 2016).
-
Each group of experiments was replicated at least three times. A histogram shows the results of the statistical analyses. Result of the experiments were evaluated by Student’s t test or F test (one-way ANOVA), and P values less than 0.05 were considered statistically significant. P less than 0.01 indicated a significant difference.
-
Microglia, immunocytes in the CNS, produce inflammatory molecules, namely NO, after stimulation (Colton, 2009). We determined the effects of HERV-W env on NO production using a NO Detection Assay Kit. HERV-W env expression was increased 3.06 fold and 1.65 fold at the mRNA and protein levels, respectively (P < 0.05, Figure 1A). A standard curve was constructed (Figure 1B). NO content increased 1.68 fold (P > 0.05), 2.52 fold (P < 0.05), 3.07 fold (P < 0.01), and 2.21 fold (P < 0.05) at 6 h, 12 h, 24 h, and 48 h, respectively, after transfection with pCMV-env. As shown in Figure 1C, NO accumulated in microglia in a time-dependent manner after overexpression.
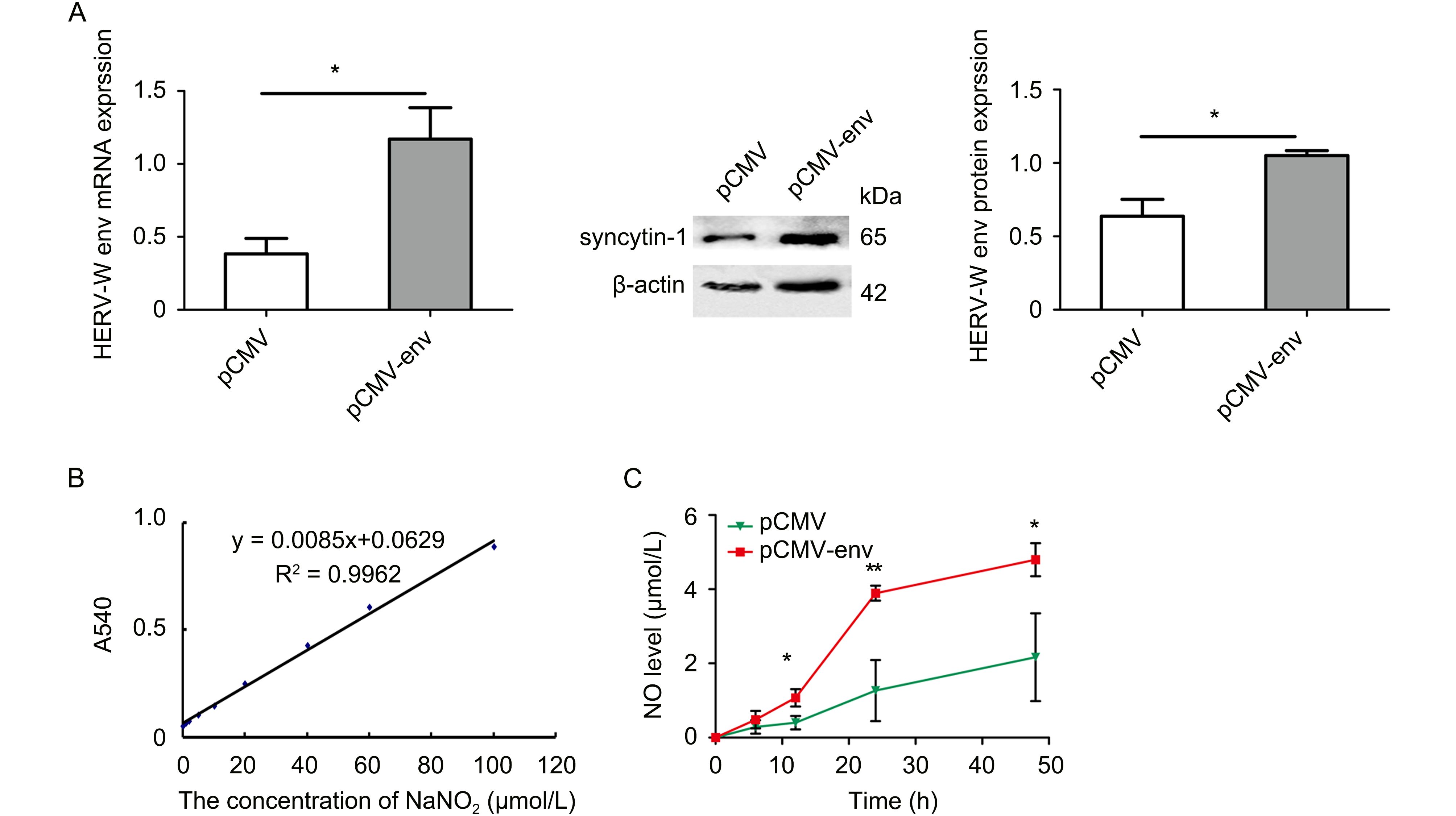
Figure 1. HERV-W env increased NO in microglia in a time-dependent manner.
-
NO is synthesized in the brain from two kinds of NOS (Kumar et al., 2017). To verify whether the increase in NO was caused by an increase in iNOS expression, real-time PCR and western blotting were used to examine iNOS expression. iNOS mRNA increased 2.55 fold in CHME-5 cells, as determined by real-time PCR (Figure 2A, P < 0.01). iNOS protein was evaluated by western blotting and was found to increase by 52.9% after transfection of CHME-5 cells with HERV-W env (Figure 2B–2C, P < 0.05). Taken together, these results indicate that HERV-W env increases expression of both iNOS mRNA and protein in microglia.
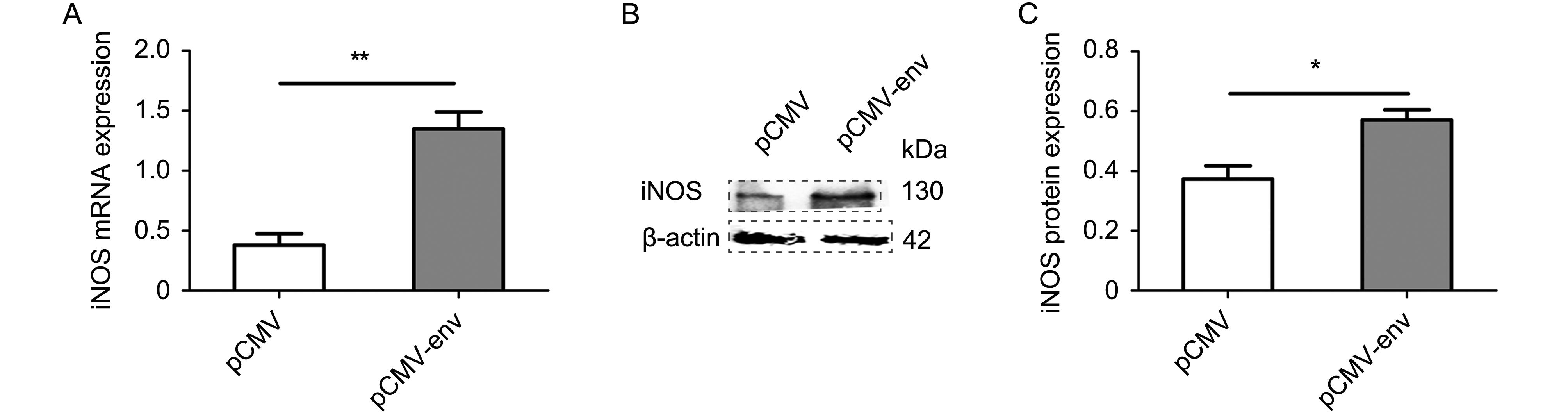
Figure 2. HERV-W env increased iNOS expression in microglia.
-
To further define the relationship between iNOS activity and HERV-W env expression, we determined the effects of HERV-W env on iNOS activity using a Nitric Oxide Synthase Assay Kit. The role of HERV-W env was confirmed by flow cytometric analysis; the fluores- cence intensity of DAF-FM DA of different groups was determined (Figure 3A–3F). As shown in Figure 3G, fluorescence intensity increased by 73% without SMT compared to with pCMV (P < 0.001), and RFU decreased by 7.1% (P > 0.05), 18% (P < 0.05), and 49% (P < 0.001) with different concentrations of SMT. These data strongly supported the idea that HERV-W env overexpression induces the activation of iNOS in microglia.
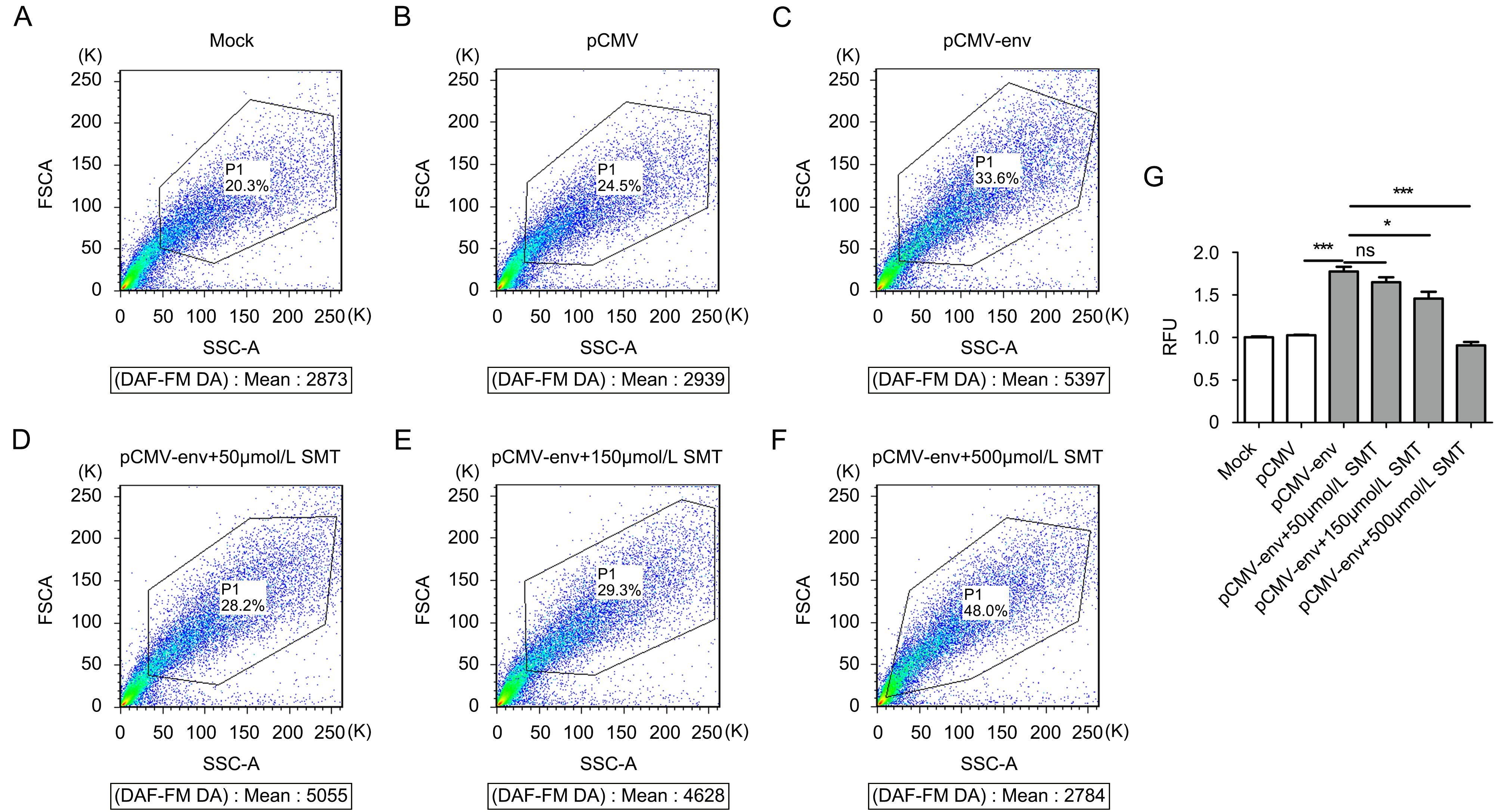
Figure 3. HERV-W env overexpression activated iNOS in microglia.
-
The iNOS promoter plays important roles in its expression (Ghasemi et al., 2014). To examine whether HERV-W env had an effect on iNOS promoter activity, cells were transiently transfected with the iNOS promoter/luciferase reporter, with or without overexpression of HERV-W env. The truncated promoters were missing segments at the 5′-terminus (Figure 4A). As shown in Figure 4B, iNOS promoter activity increased after HERV-W env transfection, with approximately 1.69-fold (P < 0.01), 1.71-fold (P < 0.01), 1.22-fold (P < 0.05), 1.25-fold (P < 0.05), and 1.44-fold (P < 0.05) increases in iNOS promoter activity observed after transfection with plasmids containing promoters 8 kb, 7 kb, 6 kb, 4 kb, and 3 kb in length, respectively. However, the 2-kb (P > 0.05) and 1-kb (P > 0.05) promoters had no significant effect on iNOS promoter activity. These results suggested that HERV-W env enhanced iNOS activation in microglia and that the 3-kb sequence of the 3′ terminus of the full-length promoter was the minimum sequence required.
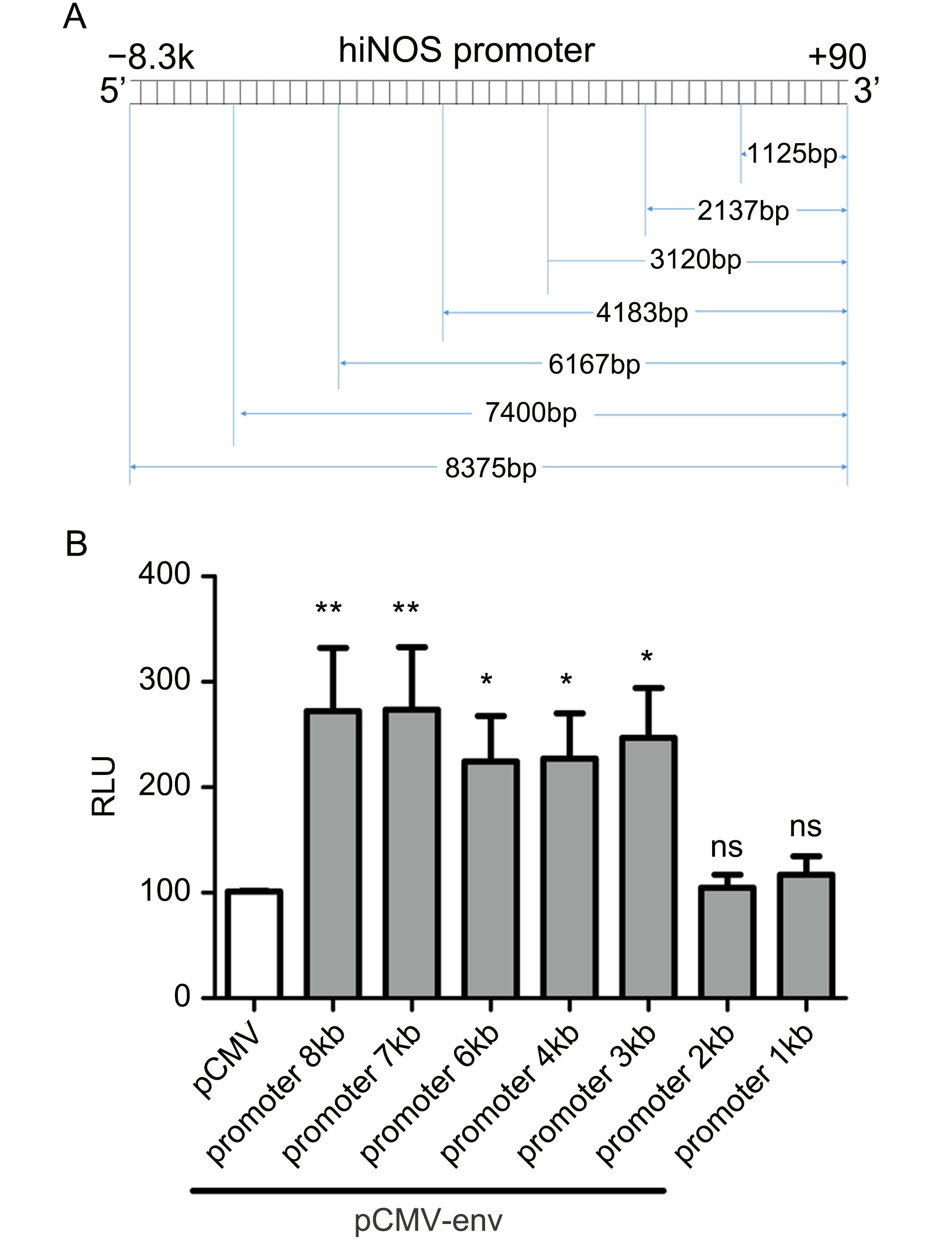
Figure 4. HERV-W env expression enhanced iNOS promoter activation.
-
Microglia migrate after stimulation such as brain injury and immunological reactions (Zhu et al., 2016). NO is a primary cause for cellular migration, and HERV-W env was previously found to increase NO. Therefore, we speculated that HERV-W env might lead to migration of microglia. To confirm the relationship between HERV-W env overexpression and microglia migration, after transfection with pCMV-env for 24 h, CHME-5 cells were transferred into a transwell chamber and cultured for another 12 h. Results showed that the number of migrated cell numbers increased after transfection with pCMV-env. The number of migrating cells with the blank vector control (pCMV) and expression vector (pCMV-env) was 10 ± 2 (means ± SME, Figure 5A) and 47 ± 2 (Figure 5B), respectively. The number of migrating cells following HERV-W env activation increased 4.7 fold (Figure 5C, P < 0.001). These data showed that overexpression of HERV-W env enhanced the migration of microglia. Moreover, results from the wound healing assay showed that the mobility of microglia increased 1.93 fold after transfection with pCMV-env (Figure 5D). These results confirmed that HERV-W env overexpression enhanced the migration of microglia.

Figure 5. HERV-W env enhanced the migration of microglia.
In conclusion, HERV-W env overexpression resulted in an increase in NO over time and upregulated the expression, function, and promoter activity of iNOS, thereby increasing the migration of microglia. These results suggest that HERV-W env expression is associated with inflammation of microglia.
-
Many copies of endogenous retroviral elements integrate into the human genome, representing about 8% of the human genome (Jern et al., 2004). Most sequences are defective (Brodsky et al., 1993). A minority of sequences are full-length copies of retroviruses (Brodsky et al., 1993) which may have the potential to be packaged into complete viral particles in the human body (Seifarth et al., 1998). HERV-W, an endogenous retroviral element, has a typical HERV structure, which is: 5′-LTR-gap-(pro)-pol-env-3′-LTR. HERV-W env is expressed in human placenta (Mi et al., 2000) and facilitates cell-cell fusion during trophoblast development (Frendo et al., 2003; Mallet et al., 2004). However, HERV-W env also contributes to various diseases, including schizophrenia (Karlsson et al., 2004), MS (Clausen, 2003), rheumatoid arthritis (Nakagawa et al., 1997), diabetes mellitus (Conrad et al., 1997), and several types of cancer (Bjerregaard et al., 2006; Gimenez et al., 2010; Wang-Johanning et al., 2007; Yu et al., 2014). These diseases are associated with inflammatory reactions. Therefore, there is a possibility that HERV-W env is activated in response to host immunity.
Because the BBB normally restricts the passage of immunocytes from the blood into the brain, the brain has been considered an "immunologically privileged site." However, immunological reactions, especially brain inflammation, occur in the CNS. NO produced by microglia plays a major role in brain inflammation. NO participates in the suppression of neutrophil adhesion and migration in acute inflammation (Adachi et al., 2000; Lefer et al., 1999).
On one hand, NO is a signaling molecule in the CNS, and, at appropriate levels, it maintains a dynamic balance (Kumar et al., 2017). On the other hand, NO freely diffusing across cellular membranes plays an essential role as a neurotransmitter in the CNS, unlike other classical neurotransmitters (Asiimwe et al., 2016). For example, NO directly modifies intracellular targets in neurotransmitter signaling pathways by crossing cellular membranes through passive transport (Mustafa et al., 2009). NO has been found to modulate physiological responses in diseases in three ways: a. stimulating host immune defenses, b. regulating neural transmissions in the brain, and c. enhancing inflammation (Jaffrey et al., 1995). A previous study implicates NO produced by microglia in the pathophysiology of a wide variety of neuropsychological diseases such as MS and AD (Scheiblich et al., 2014; Yun et al., 1997). In this study, we examined changes in NO levels when HERV-W env was overexpressed in microglia. HERV-W env was found to trigger a time-dependent increase in NO levels (Figure 1).
NO is synthesized in the body from three kinds of NOS (Kumar et al., 2017). The release of endothelial NOS (eNOS)-derived NO occurs in response to vascular permeability and angiogenesis in chronic inflammation (Wallace, 2005). NO can act as a “double-edged sword, ” which might be exploited to develop therapies for neuropsychological disorders (Dzoljic et al., 2015). iNOS, the third isoform, is induced by acute inflammation (Alderton et al., 2001). Microglia do not express iNOS in a normal physiological state, but they an express iNOS with pathologies such as inflammatory injury (Galea et al., 1992). In this study, we showed that HERV-W env upregulated iNOS expression (Figure 2). Based on previous experiments, we speculated that increased NO was a result of elevated iNOS expression. The release of high amounts of NO synthesized from iNOS is responsible for the adverse effects of NO in inflammatory sites.
Specific enzymatic activity, namely iNOS activity, is induced by stimuli and plays an important role in inflammation. Total enzymatic activity, which includes iNOS activity, has biological effects in the body. Isothiourea analogues inhibit iNOS activity by competing for iNOS binding sites on L-arginine. SMT is an isothiourea analogue. As a non-acid L-arginine analogue, SMT is a powerful and specific selective inhibitor of iNOS in humans (Alderton et al., 2001). In this study, SMT was used to inhibit iNOS to determine whether increased NO production resulted from an increase in iNOS activity. HERV-W env was found to enhance the activity of iNOS (Figure 3). Such isothiourea analogues can be employed to study of NOS isoenzymes and iNOS changes in various pathophysiological conditions.
The iNOS promoter plays an important role in regulating iNOS expression. Recently, HERV-W env expression has been proposed to increase the activation of promoters of some genes and to enhance expression of these genes in individuals such as those with schizophrenia (Huang et al., 2011; Li et al., 2013; Qin et al., 2016). Functional promoter elements of hiNOS are located upstream of the 3.8-kb and 7.2-kb promoter regions of the hiNOS gene when induced by cytokines (de Vera et al., 1996). In addition, although hiNOS uses the same cytokines to regulate transcription, hiNOS utilizes different models of gene regulation in human glial cells (Saha et al., 2006a). Thus, we examined the activity of the iNOS promoter by co-transfection with reporter plasmids and pCMV-env (Figure 4). Results showed that HERV-W env enhanced iNOS promoter activity and that the truncated 3-kb promoter sequence was necessary. Moreover, the functional and necessary promoter element that controlled hiNOS expression induced by HERV-W env was found upstream of the 3.1-kb promoter region of the iNOS gene in humans. NF-κB and MAPK signaling were found to regulate iNOS expression by regulating the activity of the hiNOS promoter. LPS mainly induces iNOS expression through the classical NF-κB pathway, whereas expression through the MAPK pathway has been identified in neuropsychological disorders (Ko et al., 2013; Li et al., 2013; Saha et al., 2006b). Therefore, we speculated that HERV-W env could affect hiNOS promoter activity through the classical NF-κB pathway and/or MAPK signaling pathway.
NO, an inflammatory molecule, promotes immunocytes to migrate to lesions by dispersing through the microenvironment. NO either stimulates or inhibits cellular migration, depending on the conditions, tissue location, and types of cells present (Fuseler et al., 2016). NO has been shown to be either a requirement or a stimulus for cellular migration. Studies have shown that NO can increase the migration of microglia cells (BV-2) and umbilical vein endothelial cells (HUVECs), and that iNOS inhibitor suppresses the migration of these cells (Scheiblich et al., 2014; Zhu et al., 2016). However, NO also inhibits the migration of tumor cells such as HepG2 and gastric cancer cells (Sang et al., 2011; Zhou et al., 2016). Therefore, NO has highly variable effects on cell migration and its presence is required for various cellular functions (Lu et al., 2015). We assessed microglia migration using Transwell and wound healing assays when HERV-W env was overexpressed (Figure 5). The results showed that NO increased with overexpression of HERV-W env, enhancing the capacity of microglial CHME-5 cells to migrate.
In this study, the data indicated that activating the expression of HERV-W env affected inflammation by causing NO to accumulate, leading to the expression of iNOS, upregulating iNOS promoter activity, and facilitating migration of microglia. In conclusion, we found that HERV-W env overexpression was associated with iNOS-derived NO overproduction in microglia and promoted inflammation of microglia. The expression of iNOS contributes to the pathogenesis of the neuropsychological disorders. Our study suggests therapeutic approaches based on reducing NO overproduction to attenuate neuropsychological diseases such as schizophrenia and MS.
-
This work was supported by grants from the National Natural Sciences Foundation of China (No. 31470264, No. 81271820, No. 30870789, and No. 30300117), the Key Program of Natural Science Foundation of Hubei Province of China (No. 2014CFA078), the Stanley Foundation from the Stanley Medical Research Institute (SMRI), USA (No. 06R-1366), to Dr. Fan Zhu and the Scientific Innovation Team Project of Hubei Province of China (No. 2015CFA009).
-
The authors declare that they have no conflicts of interest. This article does not contain any studies with human or animal subjects performed by any of the authors.
-
FZ conceived the experiments. RX, SL, XLW, QJY, and XNT carried out the experiments. RX and QC analyzed the data. YZ contributed materials and provided luciferase reporter gene plasmids containing the iNOS promoter. RX prepared the figures and/or tables and drafted the manuscript. FZ finalized the manuscript. All authors read and approved the final manuscript.
Human endogenous retrovirus W env increases nitric oxide production and enhances the migration ability of microglia by regulating the expression of inducible nitric oxide synthase
- Ran Xiao 1 ,
- Shan Li 2 ,
- Qian Cao 3 ,
- Xiuling Wang 1 ,
- Qiujin Yan 1 ,
- Xiaoning Tu 1 ,
- Ying Zhu 4 ,
-
Fan Zhu
1,5,,

- Received Date: 13 April 2017
- Accepted Date: 26 May 2017
- Published Date: 23 June 2017
Abstract: Human endogenous retrovirus W env (HERV-W env) plays a critical role in many neuropsychological diseases such as schizophrenia and multiple sclerosis (MS).These diseases are accompanied by immunological reactions in the central nervous system (CNS).Microglia are important immunocytes in brain inflammation that can produce a gasotransmitter-nitric oxide (NO).NO not only plays a role in the function of neuronal cells but also participates in the pathogenesis of various neuropsychological diseases.In this study,we reported increased NO production in CHME-5 microglia cells after they were transfected with HERV-W env.Moreover,HERV-W env increased the expression and function of human inducible nitric oxide synthase (hiNOS) and enhanced the promoter activity of hiNOS.Microglial migration was also enhanced.These data revealed that HERV-W env might contribute to increase NO production and microglial migration ability in neuropsychological disorders by regulating the expression of inducible NOS.Results from this study might lead to the identification of novel targets for the treatment of neuropsychological diseases,including neuroinflammatory diseases,stroke,and neurodegenerative diseases.

 DownLoad:
DownLoad: