










-
Dear Editor,
Here we explored the contribution of atlastin-1 (ATL1) to HIV replication for the first time. HIV-1 encodes only 15 proteins (Ayinde et al., 2010) and thus must exploit multiple host cell functions for successful infection. For example, HIV-1 envelope (Env) glycoproteins are synthesized as a polyprotein precursor, known as gp160, in the endoplasmic reticulum (ER) (Checkley et al., 2011) . Monomers of gp160 oligomerize in the ER into predominantly trimers. This oligomerization process is thought to facilitate the trafficking of gp160 to the Golgi complex. In the Golgi, gp160 is proteolytically cleaved by cellular furin or furin-like proteases at a highly conserved K/R-X-K/R-R motif, to yield the mature surface (SU) glycoprotein gp120 and the transmembrane (TM) glycoprotein gp41 (McCune et al., 1988; Hallenberger et al., 1992). Proteolytic processing of gp160 is absolutely required for activation of the fusogenic activity of Env and is thus essential for viral infectivity. Recent work has implicated the membrane-bound atlastin GTPases in the homotypic fusion of ER tubules (Hu et al., 2009; Orso et al., 2009). These proteins belong to the dynamin family of GTPases (Zhao et al., 2001) and are required for Golgi apparatus and ER morphogenesis (Rismanchi et al., 2008). ATL1 has been implicated in intracellular membrane trafficking, particular at the ER-to-Golgi interface (Zhu et al., 2003; Namekawa et al., 2007) . Therefore, we are interested in whether ATL1 is involved in viral replication through the membrane system.
To study the relationship between HIV-1 and host protein ATL1, we first examined if HIV-1 infection of HeLa-derived TZM-bl cells (which express endogenous CXCR4, transgenic CD4, and CCR5, and an integrated Tat-dependent β-galactosidase reporter gene) can influence the expression of ATL1 mRNA. We infected the TZM-bl with the X4-tropic HIV-1 strain NL4-3 (HIV-1 NL4-3) (0.1 MOI) for a period of 96 h, and quantified the mRNA levels of ATL1 by qRT-PCR. We observed that HIV-1 infection up-regulated the expression of ATL1 mRNA (Figure 1A). For further study, we constructed the ATL1 expression plasmid and named pcDNA3.1-ATL1. The gene of ATL1 (pGW1-ATL1, was supported by Junjie Hu (Hu et al., 2009) was amplified and inserted into the expression plasmid pcDNA3.1 (+) by restriction endonuclease digestion with EcoR V and Xho I. SDS-PAGE analysis of the expressed product ATL1 revealed a specific band at approximately 63 kDa after transfection pcDNA3.1-ATL1, which was stronger than the control (Figure 1B). These results indicated that ATL1 was successfully expressed. To evaluate the effect of ATL1 up-regulation on HIV-1 replication, the TZM-bl cell populations described above were transfected with pcDNA3.1-ATL1 or pcDNA3.1 (+). After 16 hours, the TZM-bl cells were infected with HIV-1 NL4-3 (0.1 MOI) and viral replication was measured on days 2 and 4 by determination of TCID 50. Up-regulation of ATL1 promoted the viral replication (Figure 1C). We noted that the up-regulation of ATL1 contributes to HIV-1 replication in infected cells, suggesting that HIV infection likely up-regulates the ATL1 and therefore exploit ATL1 for HIV-1 replication.
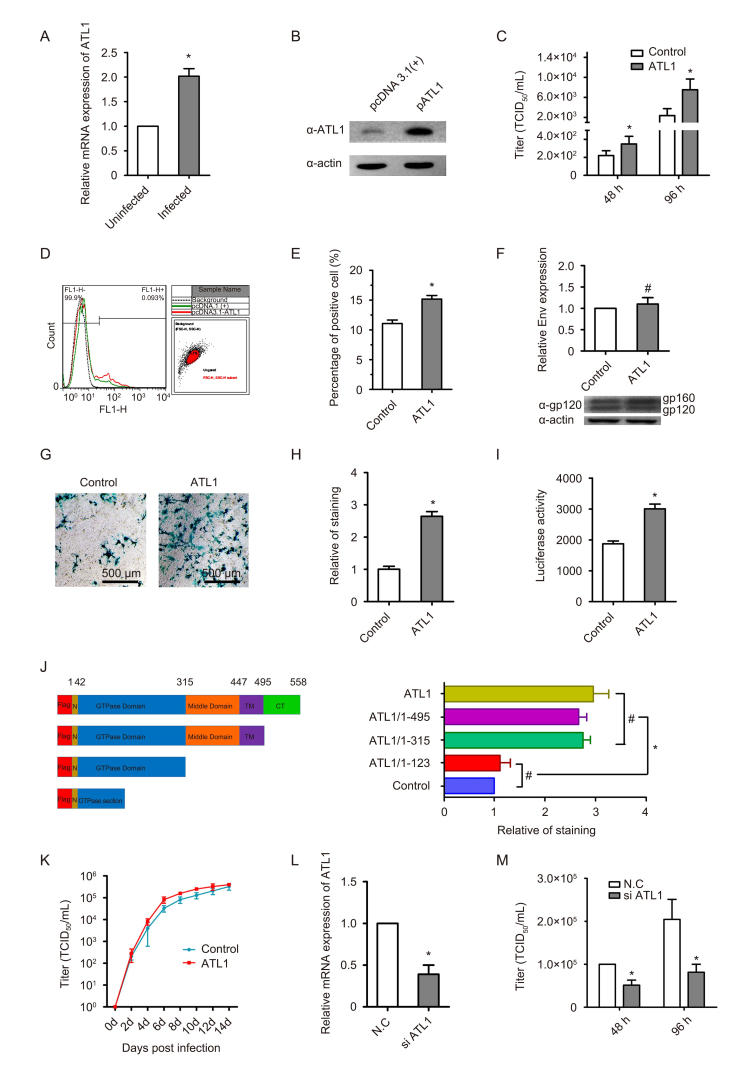
Figure 1. ATL1 promotes HIV-1 replication. (A) TZM-bl Cells were infected with HIV-1NL4-3 (0.1 MOI) for a period of 96h, and the mRNA levels of ATL1 were quantified by qRT-PCR. (B) SDS-PAGE analysis of the expressed product ATL1, after transfected with pcDNA3.1-ATL1 and transfected with pcDNA3.1 (+) as a control. ATL1 band at approximately 63 kDa. (C) The viral titers on days 2 and 4 by determination of TCID50. ATL1 expression plasmid or vector was transfected with TZM-bl cells for 16h, and then infected with HIV-1NL4-3 (0.1 MOI). Viral replication was measured on days 2 and 4 by determination of TCID50. (D, E) Surface expression level of Env. (F) Expression of total Env in cell lysates. The cell surface expression levels of envelope proteins for ATL1 or vector transfected COS-1 cells were determined by flow cytometry using anti-gp120 antibody. (G, H) COS-1 cells expressing Env were analyzed in cell-cell fusion assays by X-gal and (I) luciferase activity. (J) Cell-to-cell fusion analysis by X-gal. (Left) Schematic representation of flag tagged ATL1 or ATL1-truncated expressing plasmids. (Right) HeLa cells were co-transfected with plasmids expressing WT ATL1 or ATL1 partial fragment and pSRHS, 24 h later, cell-to-cell fusion were analyzed by X-gal. (K) HIV-1 growth kinetics in TZM-bl cells. The TZM-bl cells were transfected with pcDNA3.1-ATL1 or pcDNA3.1 (+). After 16 hours, the TZM-bl cells were infected with HIV-1NL4-3 (0.1 MOI) and viral titer was measured on days 2 to 14 by determination of TCID50. Results are representative of 3 independent experiments. #P>0.05, *P<0.05, Student’s t-test. Scale bar, 500 μm. (L) The TZM-bl cells were transfected with negative control siRNA (N.C) or ATL1 siRNA (si ATL1). The mRNA levels of endogenous ATL1 were quantified by qRT-PCR, after transfection for three days. (M) The TZM-bl cells were transfected with N.C/ATL1 siRNA. After infection, the viral titer was measured on days 2 and 4 by determination of TCID50
Since ATL1 involved in intracellular membrane trafficking, particular at the ER-to-Golgi interface, we hypothesized that the ATL1 was involved in the intracellular trafficking of Env. To test this possibility, we investigated the expression levels of Env on the cell surface, by Flow cytometric analysis. To this study, the COS-1 cell populations were transfected with pcDNA3.1-ATL1 or pcDNA3.1 (+) and Env expression plasmid pSRHS for 36 hours. For Env staining, infected cells were stained with anti-human Env primary Abs 2G12 for 30 min at 4 °C. The antibody was followed by incubation with an Alexa Fluor 488-conjugated secondary anti-human IgG antibody (Abcam). Mock (Background) -transfected cells stained in parallel were used as control. Stained cells were fixed with 1% PFA and acquired using a FACS-Calibur instrument (BD Bioscience). FlowJo was used to analyze and quantify the data. Anti-HIV gp120 Abs 2G12 were obtained through the NIH AIDS Research and Reference Reagent Program from Dr. Hermann Katinger ( Trkola et al., 1996). Surface expression studies revealed that the ATL1 can promote the level of Env expression on the cell surface, because the level of Env expression of ATL1-transfected cells is higher than the control (Figure 1D, 1E). We analyzed the protein profiles of total Env by immunoblotting, using anti-gp120 Abs, we found that the difference in total Env expression against control and ATL1-transfected cells was not statistically significant (Figure 1F).
We next assessed if ATL1 is involved in the spread of viral progeny in adjacent cells. To this purpose, we investigated whether ATL1 regulates the Env-mediated cell-to-cell fusion. The assay for cell-cell fusion mediated by Env has been described previously (Bhakta et al., 2011). COS-1 cells (4 × 10 4) were transfected with 300 ng HIV-1 Env expressing plasmid pSRHS (Salzwedel et al., 1993), together with 500 ng ATL1 expression vectors pcDNA3.1-ATL1 or with 500ng vectors pcDNA3.1(+), in 24-well plates using polyethyleneimine (PEI) (Sigma-Aldrich, MO, USA). After 24 h, the transfection cells were co-cultured with TZM-bl cells (1.2 × 10 5) for another 24 h. For X-gal staining, cells were washed with PBS and fixed in 0.8% formaldehyde for 5 min and then stained for β-galactosidase activity with X-gal (Sangon, Shanghai, China). Blue-stained foci were visualized under 100× magnification. For the luciferase assay, cells were rinsed with PBS, disrupted with lysis buffer, and centrifuged at 13, 200 × g for 3 min. The clarified lysated (20 μL) was mixed with luciferin reagent and luciferase activity measured using the Steady-Glo Luciferase Assay System (Promega, WI, USA). The results showed that ATL1 can promote the cell-to-cell fusion (Figure 1G, 1H, 1I). We interesting found that the enhancement for Env expression in Figure 1D, 1E, 1F, is modest, while the contribution to cell-to-cell fusion is significant (Figure 1G, 1H, 1I). The reasons for this phenomenon may have the following two aspects. The first reason is that slightly enhanced Env might effectively promote the cell-to-cell fusion. The second reason is that the amount of the functional Env at cell surface does not have linear relation with the degree of X-gal staining in the cell to cell fusion assay. A donor cell possibly fuse with one or more target cells, so the cell-to-cell fusion experiment is more significant than the Env expression experiment.
Previous studies have been reported that the ATL1 consisted of the GTPase domain, two closely spaced transmembrane (TM) segments, middle domain and cytoplasmic domain (CT) (Figure 1J) (Bian et al., 2011).
Next, we constructed a series of C-terminal-truncation plasmids to define the key domain required for the ATL1 in the functional roles of promoting the cell-to-cell fusion. The amino acid sequences and nomenclature of the ATL1 truncation mutants are shown in Figure 1J (left). Truncation fragments were amplified and inserted into the expression plasmid pCMV-Tag2b by restriction endonuclease digestion with Hind Ш and BamH I. We performed the X-gal method to detect the Env-mediated cell-to-cell fusion. As shown in Figure 1J (right), with the exception of ATL1/1-123 and control, we observed similar fold change of the cell-to-cell fusion with ATL1 WT or ATL1 fragments. All the results indicate that the GTPase domain of ATL1 is the key domain of promoting Env-regulated cell-cell fusion.
To know the impact of ATL on the virus replication cycle, we assessed the HIV-1 growth kinetics in TZM-bl cells. The TZM-bl cells were transfected with pcDNA3.1-ATL1 or pcDNA3.1 (+). After 16 hours, the TZM-bl cells were infected with HIV-1 NL4-3 (0.1 MOI) and viral titer was measured on days 2 to 14 by determination of TCID 50. Through the growth kinetics, we found that the titer in ATL1-transfected cells was higher than the titer in vector-transfected cells and with statistically significance (Figure 1K).We found that ATL1 overexpression can apparently promote virus replication in the first 6 days; After 8 days, the titer of the control group was gradually equal to the titer of the ATL1 expression group. We think that the reason is that the replication of the virus is close to the plateau, and the virus is getting saturated.
Finally, endogenous altastin-1 in TZM-bl cells was knock-down to confirm the contribution of atlastin-1 (ATL1) to HIV replication. The TZM-bl cells were transfected with negative control siRNA (5′-UUCUCCGAACGUGUCACGUTT-3′) or ATL1 siRNA (5′-GCGAGAGACCACAGGAAUUTT-3′; 5′-GCUACCUCAUCCUGGCUUATT-3′) at day 1 and day 3. Successfully knockdown was confirmed by qRT-PCR (Figure 1L). At the day 4, the TZM-bl cells were infected with HIV-1 NL4-3 (0.5 MOI) and viral titer was measured on days 2 and 4 after infection by determination of TCID 50. The results showed that the down-regulation of ATL1 inhibited the viral replication (Figure 1M).
All these results indicated the regulatory role of ATL1 in HIV replication. It should be pointed out that the results were exclusively based on the widely used laboratory-adapted strain NL4-3 and transformed cell lines, which is a limitation of this study. It remains a question whether HIV replication is affected by ATL1 in physiological conditions. Primary cells and other HIV-1 strains will be included in future study of the mechanism of the regulation.
In conclusion, we first found that HIV infection up-regulates the ATL1 mRNA. Up-regulation of ATL1 contributes to HIV-1 replication and down-regulation of ATL1 in host cells inhibited the viral replication. The further research found that ATL1 promoted the level of Env expression on the cell surface and then promoted the cell-to-cell fusion. Truncated mutation experiments revealed that the GTPase domain of ATL1 is the key domain of promoting Env-regulated cell-cell fusion. Finally, through the kinetic experiment, we found that ATL1 was able to promote long-term replication of HIV-1. We believe that the characterization of ATL1 contributes to a better understanding of HIV-host interactions and can identify potential novel antiviral targets.
HTML
-
This work was supported by grants from the National Natural Science Foundation of China (81371820, 81571987) and the Ph.D. Candidate Research Innovation Fund of the School of Medicine Nankai University. We greatly appreciate the gift of pGW1-ATL1 from Junjie Hu (Chinese Academy of Sciences, Beijing, China). The authors declare that they have no competing interests. This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: