














-
Dear Editor,
Clustered regularly interspaced short palindromic repeat (CRISPR)-associated system (Cas) is an adaptive immune system discovered in prokaryotic bacteria or archaea that can fend off invading nucleic acids. Because of its simplicity, high efficiency and versatility, CRISPR/Cas system-mediated genome editing has been widely applied in plant research and agricultural production. In brief, an effective CRISPR/Cas system contains an easily-engineered guide RNA (gRNA) and a Cas effector protein. Recently, a special type of RNA targeting CRISPR/Cas-associated protein, Cas13a (formerly called C2c2), was found (Shmakov et al. 2015). This new type of Cas protein, unlike Cas9 or Cas12a, can target and edit single-stranded RNA (ssRNA) rather than double-stranded DNA (dsDNA) in vivo (Abudayyeh et al. 2017; Cox et al. 2017).
Plant viruses, of which RNA viruses constitute the largest group, are obligate invading parasites that cause devastating diseases in economically important crops. There exist several RNA-targeted anti-viral immunity systems in plants, including RNA silencing, RNA decay and other RNA quality control systems (Li et al. 2019), some of which have been applied into the prevention and control of plant RNA viruses in the field, such as the effective RNA silencing-based antiviral engineering. As the development of CRISPR/Cas systems, these systems-mediated DNA or RNA editing/interference in plants make them very attractive tools applicable to inhibit the infection of different plant RNA viruses (Cao et al. 2020). Generally, targeting viral RNA sequences and editing DNA sequences of host susceptibility genes are two main CRISPR/Cas technology-mediated strategies to be used for restriction of viral infection in plants (Cao et al. 2020; Pyott et al. 2020). A plant-codon-optimized version of FnCas9 paired with rgRNA targeting plant ssRNA viruses has been employed to gain resistance to several agricultural important plant RNA viruses (Zhang T et al. 2018). Recently, the use of Cas13a as an effective method to inhibit plant RNA virus infection was also demonstrated (Zhang T et al. 2018). Eukaryotic translation initiation factors eIF4E, eIF4G and their isoforms eIF(iso)4E and eIF(iso)4G are the most widely exploited host susceptibility genes edited by CRISPR/Cas9 systems to confer resistance to plant RNA viruses (Chandrasekaran et al. 2016; Cao et al. 2020).
A recent work characterizing the functionality of Cas13d from Ruminococcus flavefaciens (CasRx) has shown that the single effector CasRx protein is a programmable RNA-guided ssRNA nuclease. CasRx possesses two higher eukaryotic and prokaryotic nucleotide binding domains (HEPN) that dictate CRISPR RNA maturation and target cleavage (Konermann et al. 2018; Zhang C et al. 2018; Zhang B et al. 2019). CasRx is smaller than Cas13a, Cas13b and other Cas13 variants, and it is effective to combat RNA virus (Mahas et al. 2019). In addition, CasRx is independent of protospacer flanking site (PFS), and HEPN-dependent ribonuclease with RNA cleavage activity (Konermann et al. 2018; Wessels et al. 2020). In this case, besides cleaving the target RNA, CasRx also shows promiscuous ability of the RNase to cleave collateral RNAs once activated in the presence of template targets. Therefore, CasRx-mediated RNA-targeting tools show promise for applications such as RNA virus suppression, gene knockdown, RNA detection, or transcript tracking. Here, we systemically analyzed the effect of CasRx-mediated RNA interference in Nicotiana benthamiana plants defending against exogenous RNAs expressed from a plant virus or from a transiently transformed vector.
The CasRx-mediated RNA interference system in N. benthamiana plants was firstly engineered in this study. The recombinant construct pCambia1300-CasRx (CasRx) was based on the pCambia1300 backbone vector, which includes the enhanced Pol Ⅲ promoter 35S (e35S) from cauliflower mosaic virus (CaMV), a plant-codon-optimized CasRx gene, and the Nos terminator (Fig. 1A). For the generation of crRNA transcripts, we used two different strategies. One single-guide RNA expression strategy used the Arabidopsis thaliana small nucleolar RNA U6 (AtU6) promoter, while the other one used a virus-based vector, tobacco rattle virus (TRV), which harbors the promoter from pea early browning virus (PEBV), and it has been proven to efficiently mediate expression of the sgRNA transcripts and work well with SpCas9 (Ali et al. 2015). It is worth mentioning that we constructed an all-in-one vector, which contains the CasRx and the AtU6-crRNA, in the AtU6 sgRNA expression strategy (Fig. 1A). In comparison to a previous report (Mahas et al. 2019), all sgRNAs are composed of 30 nt direct repeats and 22 nt spacer in this study, which is redesigned and optimized by the discover researcher of CasRx (Konermann et al. 2018).
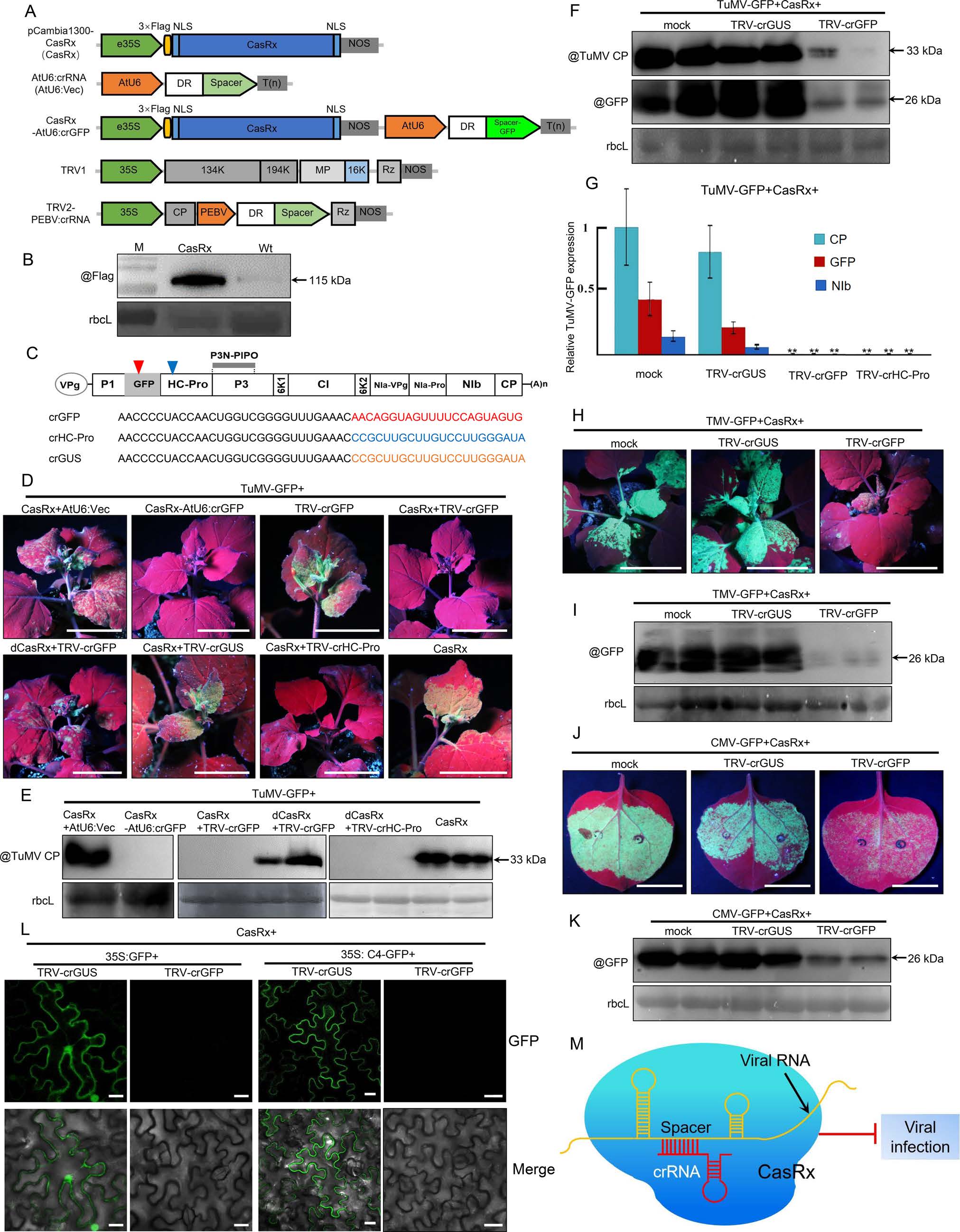
Figure 1. The CasRx-mediated suppression of plant RNA virus and RNA expressed from a transiently transformed vector. A Schematic representation of CRISPR/CasRx systems. CasRx was fused with 2 9 SV40 nuclear localization signal (NLS) and 3 9 Flag tags driven by the 35S promoter (CasRx). The target sequences can be inserted into AtU6: crRNA vector or tobacco rattle virus (TRV)-based vector. CasRx-AtU6: crGFP is a fused vector, which can express the CasRx protein driven by the 35S promoter and a specific sgRNA targeting the GFP RNA driven by AtU6 promoter. For the TRV-PEBV vector, sgRNA is engineered into the TRV2 vector driven by the pea early-browning virus (PEBV) promoter. B Western blotting analysis of the expression of CasRx using anti-Flag antibodies. Agrobacterium cultures including pCambia1300- 3 9 Flag-CasRx were infiltrated into Nicotiana benthamiana leaves, and total protein was extracted from the infiltrated leaves (CasRx) or wild-type leaves (Wt) for Western blotting analysis at 48 h post infection. Coomassie brilliant blue (CBB)-staining of RuBisCO large subunit (rbcL) serves as a loading control (B, E, F, I, K). C, Schematic diagram of turnip mosaic virus fused with GFP (TuMV-GFP). The red or blue triangle indicates the crRNA targeting sequences to GFP or HCPro, and the sequences of these crRNA (crGFP, crHC-Pro and crGUS) are listed here. D, GFP fluorescence was photographed from the systemic leaves infected with TuMV-GFP and other indicated components. Agrobacterium cultures harboring the following constructs to express TuMV-GFP together with CasRx and AtU6:Vec, or TuMV-GFP together with CasRx-AtU6:crGFP, or TuMV-GFP together with TRV1 and TRV2-PEBV: crGFP (TRV-crGFP)/TRV2-PEBV: crGUS (TRVcrGUS)/TRV2-PEBV: crHC-Pro (TRV-crHC-Pro), or TuMV-GFP together with CasRx and TRV-crGFP, or TuMV-GFP together with dCasRx and TRV-crGFP were co-infiltrated into N. benthamiana leaves, and the infiltrated plants were photographed under UV light at 7 dpi. Scale bar = 4 cm. E, F, Immunoblotting analysis of total protein extracted from upper non-inoculated leaves of plants indicated in D. Infiltration buffer was used as mock. Anti-TuMV coat protein (CP) and anti-GFP antibodies were used in these assays. Two independent experiments were performed; representative results are shown (D–F). G, Relative expression level of the indicated genes in the systemic leaves infected by TuMV-GFP, CasRx together with mock (infiltration buffer), or TRV-crGUS, or TRV-crGFP, or TRV-crHC-Pro at 7 dpi. The relative TuMV CP, GFP, or TuMV NIb RNA levels were analyzed by RTqPCR, and NbActin served as an internal reference gene. These data were analyzed using two-sided Student's t-test, **P < 0.01. H, GFP fluorescence was photographed from the plants infected with TMV-GFP and other indicated components. Agrobacterium cultures harboring constructs to produce TMV-GFP, CasRx together with mock, or TRVcrGUS, or TRV-crGFP were co-infiltrated into N. benthamiana leaves, and the infected plants were photographed under UV light at 7 dpi. Scale bar = 4 cm. I, Immunoblotting analysis of total protein extracted from the upper non-inoculated leaves of plants indicated in H using anti-GFP antibodies. J, Visualization of GFP fluorescence in the infiltrated leaves inoculated with CMV-GFP and other indicated components at 5 dpi. Scale bar = 5.5 cm. K, Immunoblotting analysis of total protein extracted from the infiltrated leaves indicated in J using anti-GFP antibodies. L, Confocal images of the infiltrated leaf patches coexpressing 35S: GFP or 35S: C4-GFP together with TRV-crGUS or TRV-crGFP. Three independent experiments were performed and 20 cells were observed each time; representative images were shown. Scale bar: 20 μm. M, A working model for the CasRx system-mediated suppression of the invading viral RNA. CRISPR/CasRx systemmediated viral RNA cleavage leads to the suppression of the plant virus infection. Therefore, we propose that the CRISPR/CasRx system can be engineered into plants to achieve viral resistance.
The efficient expression of CasRx in N. benthamiana leaves by agro-infiltration was confirmed by Western blotting using anti-Flag antibodies (Fig. 1B). Turnip mosaic virus (TuMV) is a single-stranded, positive RNA (+ ssRNA) virus belonging to potyviruses, which infects a broad range of host plants and causes a devastating disease in the field. Here, we utilized TuMV to evaluate whether CRISPR/CasRx-mediated RNA interference can attenuate RNA virus infection in plants. The infectious clone TuMV-GFP, which can be tracked by GFP fluorescence under the UV lamp was used in this study (Fig. 1C). The CasRx RNase cleavage activity strictly relies on its HEPN motif, which contains the highly conserved R-X4-H residues in the family of Cas13 homologues (Konermann et al. 2018; Zhang C et al. 2018; Zhang B et al. 2019). To test whether these residues are required for the CasRxmediated anti-viral activity, we generated a mutant version of the protein by replacing the key specific residue motif R-X4-H with A-X4-A (dCasRx). Agrobacterium cultures harboring the following constructs to express TuMV-GFP, CasRx, and AtU6:crRNA (AtU6:Vec), or TuMV-GFP and CasRxAtU6:crGFP, or TuMV-GFP, TRV1 and TRV2-PEBV: crGFP (TRV-crGFP), or TuMV-GFP, CasRx and TRV-crGFP, or TuMV-GFP, dCasRx and TRV-crGFP, or TuMV-GFP, CasRx, TRV1 and TRV2-PEBV: crGUS (TRV-crGUS), or TuMV-GFP, CasRx, TRV1 and TRV2-PEBV: crHC-Pro (TRV-crHC-Pro), or TuMV-GFP and CasRx were mixed together before infiltration into the leaves of N. benthamiana plants (5–6 leaf stage) to test its suppression effect on viral infection. As shown in Fig. 1D–1F, the expression of crGFP driven either by the AtU6 or the PEBV promoter together with CasRx, or the expression of crHC-Pro driven by the PEBV promoter together with CasRx could achieve efficient suppression on TuMV-GFP systemic infection. Compared to the control (AtU6:Vec or TRV-crGUS), the expression of crRNA from TuMV-GFP sequences significantly decreased the accumulation of viral coat protein (CP) or the GFP protein in the systemic leaves at 7 days post infiltration (dpi). In contrast, dCasRx failed to inhibit TuMV-GFP infection and did not affect the GFP protein accumulation, indicating that these effects require CasRx RNase activity (Fig. 1D–1E). In addition, expression of TRV-crGFP without CasRx was unable to inhibit TuMV-GFP infection and viral protein accumulation, suggesting that this system is dependent on CasRx, rather than virus-induced RNA silencing (Fig. 1D–1F). Consistent with these observations, RT-qPCR showed that the accumulation of GFP RNA, TuMV NIb RNA, and TuMV CP RNA was significantly decreased in the presence of the functional CasRx system duplexed with crRNA targeting GFP or HC-Pro (Fig. 1G). Furthermore, we also sequentially inoculated TRV vector-based programmable CasRx systems (CasRx and TRV-crGUS, TRV-crHC-Pro or TRV-crGFP) prior to the TuMV infectious clone to check the CasRx-mediated resistance. As shown in Supplementary Fig. S1, the sequential inoculations of the programmable CasRx systems prior to TuMV also exhibited the significant suppression on TuMV infection and viral RNA accumulations. Collectively, our data show that this CasRx-mediated anti-viral system could attenuate viral infection by decreasing viral RNA and protein accumulation, in which the active CasRx protein, the generation of mature crRNA, and the target ssRNA virus are indispensable for its efficient activity.
To explore the universal application of CRISPR/CasRx, we tested this system on other plant RNA viruses from different families, namely tobacco mosaic virus (TMV) and cucumber mosaic virus (CMV). As expected, the expression of the CasRx duplexed with the sgRNA targeting GFP driven by the PEBV promoter in the TRV2 vector was able to remarkably attenuate the systemic infection of TMV-GFP or the local CMV-GFP expression and decrease the GFP protein accumulation (Fig. 1H–1K). Of note, CMV CP was replaced with GFP in this vector, so the resulting virus is unable to move onto systemic leaves. These results suggest that the CasRx-mediated anti-viral system represents an efficient tool to inhibit the infection of an array of viruses by targeting the conserved region of the viral genomic RNA. In addition, we also examined whether the CasRx-mediated RNA interference was capable of inhibiting the transcribed RNA from a transiently transformed vector. As shown in Fig. 1L, the CasRx-mediated GFP interference could efficiently inhibit the expression of GFP or C4-GFP (tomato yellow leaf curl C4 protein fused with GFP).
In summary, we have established a CasRx-mediated RNA interference system which is capable of attenuating plant RNA virus infection and suppressing the expression of the target RNA. Our results greatly expand the applicability of CasRx and provide a new tool for the design of anti-viral strategies as well as for RNA functional studies in plants (Fig. 1M).
HTML
-
This work is funded by the National Natural Science Foundation of China (31972244 and 31930089). Apologies to colleagues whose work could not be discussed due to space limitations.
-
The authors declare that they have no conflict of interest.

 DownLoad:
DownLoad: