














HTML
-
Transcription of human immunodeficiency virus type 1 (HIV-1) is regulated by distinct viral and cellular transcription factors that often bind to viral long terminal repeat (LTR) [review in (Karn and Stoltzfus 2012)]. Transcription silencing of the LTR often drives the proviral DNA into latency, thus creating HIV-1 reservoirs, a major barrier to HIV-1 eradication (Shang et al. 2015; Khoury et al. 2018; Delannoy et al. 2019). Either the absence of key cellular transcription activators such as NF-jB or the presence of strong transcription suppressors can lead to the shutdown of HIV-1 LTR promoter. The identification of these cellular factors and characterization of their mechanistic action often help the search of therapeutic agents aiming at HIV-1 cure.
The cyclic AMP-responsive element (CRE)-binding protein (CREB) regulates the expression of genes that contain CRE at their promoters. The action of CREB responds to a wide variety of signals, including hormonal and nutrient cues (review in Bonni et al. 1995; Altarejos and Montminy 2011). The cAMP-regulated transcriptional co-activators (CRTCs) were first identified as co-factors of CREB, associating with CREB and increasing CREB activity (Altarejos and Montminy 2011). The CRTC family has three members, CRTC1, CRTC2 and CRTC3. They share similar structures, an N-terminal CREB-binding domain (CBD), a central regulatory (REG) domain, a splicing domain (SD) and a C-terminal TAD (Conkright et al. 2003; Iourgenko et al. 2003). CRTC1 is expressed predominantly in the brain, CRTC2 and CRTC3 are ubiquitously expressed (Conkright et al. 2003; Uebi et al. 2010; Watts et al. 2011). CRTC2, the most extensively studied member of the CRTC family, is a nucleocytoplasmic shuttling protein, and regulates multiple cellular signaling pathways (Iourgenko et al. 2003; Watts et al. 2011). The phosphorylated CRTC2 is sequestered within the cytoplasm. In response to cAMP and calcium, CRTC2 is dephosphorylated and translocates into the nucleus, where it associates with CREB and stimulates the expression of downstream genes (Iourgenko et al. 2003; Watts et al. 2011). Beyond their role in transcription, CRTCs also modulate the alternative splicing of some CREB target genes. This latter function requires its conserved Pro-rich domain (Amelio et al. 2009).
In addition, CRTC proteins have been reported to modulate virus replication. Both CRTC1 and CRTC2 enhance hepatitis B virus (HBV) transcription and replication (Tang et al. 2014; Tian et al. 2014). All three CRTC proteins assist the activation of human T-cell leukemia virus 1 (HTLV-1) LTR promoter by viral Tax protein (Siu et al. 2006; Jiang et al. 2009). The Tax protein hijacks the phosphorylated CREB, CRTC2 and p300 to activate CREdependent Cyclin D1 transcription (Kim et al. 2010).
An early study identified a cAMP response element within HIV-1 LTR, which is bound by the AP-1 and CREB/ATF (activating transcription factor) proteins and thus allows HIV-1 to respond to cAMP/PKA (protein kinase A) activation signals (Rabbi et al. 1997). These data are further supported by the increase in HIV-1 transcription as a result of the activation of the cAMP pathway (Banerjee et al. 2011, 2017). Moreover, p300/CBP (CREB-binding protein), another co-activator of CREB, also binds to the HIV-1 LTR, stimulates HIV-1 gene expression when the viral DNA is integrated in transcriptionally silent region of the cellular DNA (Wong et al. 2005). In contrast, the role of CRTC proteins in HIV-1 gene expression remains uninvestigated.
Here, we report that HIV-1 infection induces CRTC2 expression, but CRTC2 diminishes HIV-1 production and viral RNA expression. We further observed that CRTC2 reduced RNA Pol Ⅱ occupancy at LTR and inhibited HIV-1 LTR transcription, thus suppressing HIV-1 gene expression. As a result, knockdown of CRTC2 activates latent HIV-1. These data together suggest that CRTC2 is capable of facilitating HIV-1 latency, and reveal a new host target for the development of HIV-1 cure strategies.
-
HEK293T, A549 cells, Huh7.5.1 cells and Vero E6 cells (ATCC) were were cultured in DMEM (GBICO) supplemented with 10% fetal bovine serum (FBS) (GBICO). SupT1 cells (ATCC) were maintained in RPMI-1640 (GBICO) containing 10% FBS. The latently infected cell line Jurkat E4 (CD4+ T cells harboring an HIV-1 proviral DNA encoding the GFP reporter), was kindly provided by Dr. Pengyu Chen (Tianjin Medical University). These cells were maintained in RPMI-1640 (GBICO) containing 10% FBS. Peripheral blood mononuclear cells (PBMCs) were purchased from Beijing Reader Biotechnology Co., Ltd, were isolated from the buffy coats of healthy volunteers using density gradient centrifugation on Lymphoprep. PBMCs were cultured in complete RPMI 1640 medium containing 20% fetal bovine serum, 100 mg/mL streptomycin and 100 U/mL penicillin) plus 50 lg/mL concanavalin A (TargetMol) and 1/1000 recombinant human interleukin 2 (IL-2) (R & D) for 3 days and then infected with vesicular stomatitis virus glycoprotein (VSVG)- pseudotyped HIV-1. HEK293T cells were transfected with Lipofectamine 2000 (Invitrogen), Jurkat E4 cells were transfected with the Neon transfection Kit (Invitrogen) according to the manufacturer's instructions. The cDNA sequences of CRTC1 (NM_015321), CRTC2 (NM_181715), and CRTC3 (NM_022769) were cloned into pcDNA3.1 vector (Invitrogen). They all contain the Flag tag at the C-terminus. All the CRTC2 truncations were also constructed into pcDNA3.1 vector with the Flag tag at the C-terminus. The CRTC2 point-mutations were constructed with Muta-Direct Site-Directed Mutagenesis Kit (SBS genetech Co., Ltd) according to the manufacturer's instructions.
VSV-G pseudotyped HIV-1 pNL4-3Luc(R-E-), A/WSN/33 (H1N1), JFH1 HCVcc and LINE-1 were described previously (Hu et al. 2015; Wang et al. 2019; Yi et al. 2019). Influenza A virus (strain A/Puerto Rico/8/1934 H1N1) was provided by Dr. Yuelong Shu from Chinese National Influenza Center, China; ZIKV (strain FSS13025) was provided by Dr. Mark Wainberg from McGill University, Canada.
-
Cells were lysed in buffer containing 25 mmol/L Tris, pH 7.4, 150 mmol/L NaCl, 1% NP-40, 1 mmol/L EDTA, 5% glycerol (Pierce) on ice for 1 h, then centrifuged at 12, 000 rpm for 10 min to remove cell debris. Cellular extracts were subject to electrophoresis in SDS-PAGE. Proteins were transferred onto polyvinylidene difluoride (PVDF) membranes and probed with the indicated antibodies at predetermined concentrations. Western blots were performed with antibodies that are specifically reactive with HIV-1 p24 (NIH), HA (Abcam), flag (Sigma) and b-actin (ab8224), followed by incubation with corresponding secondary antibodies. Proteins were detected with enhanced chemiluminescence (Millipore). Bands in Western blots were quantitated using ChemiDoc (Bio-Rad) automated digitizing system and Image J software.
-
ShRNA-expressing lentiviral vectors were purchased from Sigma-Aldrich (TRCN0000229833, NM_181715.1). The pLKO.1 vector containing a scrambled shRNA was used as control. Pseudotyped viral stocks were produced by transfecting HEK293T cells with shRNA-expressing lentiviral vector DNA together with the lentiviral packaging DNA constructs (Naldini et al. 1996). Levels of viruses produced were detrmined by measuring viral p24. HEK293T cells were infected with virus (1 ng of p24 per 106 cells) expressing shRNA against CRTC2 and selected with puromycin (2 mg/mL; Sigma).
-
HEK293T cells were co-transfected with HIV-1 DNA (NL4-3, BH10, or AD8) and CRTC2 plasmid. The production of nascent viruses in the supernatants was measured by infecting TZM-bl indicator cells. To assess the effect of CRTC2 on VSVG pseudotyped HIV-1, HEK293T cells were co-transfected with 300 ng pNL4-3Luc(R-E-), 200 ng VSVG and CRTC2-flag (0, 200, 500, or 1000 ng) DNA in 6-well-plate. Forty-eight hours after tranfection, supernatants were collected and filtered through a 0.45 μm filter, and then used to infect the SupT1 cells (1 × 105) in 96-well plates. Forty-eight hours later, SupT1 cells were lysed, firefly luciferase activities were determined using a firefly Luciferase Assay System (Promega).
-
In order to determine whether the CRTC2-mediated inhibition is specific to HIV-1, A549 cells, Huh7.5.1 cells and Vero E6 cells were transfected with CRTC2 cDNA plasmid, 24 h later, cells were infected with IAV-1, HCV, Zika virus or co-expressed with LINE-1 respectively. After 48 h, cell lysis was analyzed by Western blot using antibodies against NP (IAV), Core (HCV), NS3 (ZIKV) and ORF1p (LINE-1) proteins.
-
To measure the HIV-1 LTR promoter activity, HEK293T cells were transfected with HIV-1 LTR-luciferase, pRLTK, and CRTC2 DNA or empty vector. The pRL-TK vector serves as the control for transfection efficiency. Forty-eight hours post-transfection, activities of firefly luciferase (from HIV-1 LTR) and renilla luciferase from (TK promoter) were measured using the Dual-luciferase assay system (Promega). Similarly, HEK293T cells were transfected with LINE-1 UTR-luciferase and pRL-TK DNA. All luciferase data were collected on Centro XS3 LB 960 Microplate Reader and Mikrowin software (Berthold).
-
Experiments were performed using the EZ-ChIPTM Chromatin Immunoprecipitation Kit (Millipore, Catalog # 17-371) according to the manufacturer's instructions. The anti-PolII antibody (sc-9001, Santa Cruz) was used. AntiIgG (sc-2027, Santa Cruz) was used as a negative control. Immunoprecipitated DNA was analyzed by real-time PCR using the KAPA SYBR FAST qPCR Kit (TAKARA) and the Mx3000PTM Quantitative PCR System thermal cycler (Stratagene). Sequences of the ChIP primers used for different HIV-1 RNA regions are: HIV-LTR proximal region: proximal-for, 5'-GTTAGACCAGATCTGAGCCT-3', proximal-rev, 5'-GTGGGTTCCCTAGTTAGCCA-3'; HIV-LTR distal region: distal-for, 5'-TGGGAGCTCTCTGGCTA ACT-3', distal-rev, 5'- TGCTAGAGATTTTCCACACT GA-3'; CMV proximal region: proximal-for, 5'-ATATT GGCCATTAGCCATA-3', proximal-rev, 5'-TGTAACGCG GAACT CCATAT-3'; Distal (gag) regions: distal-for, 5'-TCAGACA GGATCAGAAGAAC-3', distal-rev, 5'-ACG CGTCCTGA AGCTTATG-3'. PCR was performed at 95 ℃ for 10 min, followed by 40 cycles of 95 ℃ for 30 s and 60 ℃ for 30 s. DNA enrichment was calculated as the percentage of immunoprecipitated DNA under each condition relative to corresponding DNA input [100 * ((Ct IP/Ct INPUT) - (Ct IgG/Ct INPUT)), where Ct is the cycle at which the threshold line is crossed].
-
HEK293T cells were transfected with CRTC2-flag DNA in 6-well plate, then infected by VSVG-pseudotyped HIV-1. 48 h later, total DNA was isolated using DNeasy DNA isolation kit (Qiagen), and total RNA was isolated using TRIzol (Invitrogen). Alu-LTR was performed to measure integrated provirus cDNA using primers (Alu-LTR-forward, 5'-TCCCAGCTACTCGGGAGGCTGAGG-3'; AluLTR-reverse, 5'-AGGCAAGCTTTATTGAGGCTTAAG- 30; Internal-forward, 5'-CACACACAAGGCTACTTCCC T-3'; Internal-reverse, 5'-TAGCCACTCCCCAGTCCCGC CC-3'; HIV-1 RNA was quantified with qRT-PCR with the internal primers as above. Levels of GAPDH DNA or RNA were determined as control, using primers GAPDH-forward (5'-GAAGGTGAAGGTCGGAGT-3') and GAPDHreverse (5'-GAAGATGGTGATGGGATTTC-3'). Primers for unspliced (US), single spliced (SS), and mutiple spliced (MS) HIV transcripts are employed to quantify different HIV transcripts. LTR-forward: 5'-TTGTGTGACTCTGGTAACTA-3'; Gag-reverse: 5'-GCTTAATACCGACGCTC T-3'; Vif-reverse: 5'-GTACTTTCATAGTGATGTCT-3'; Vpr-reverse: 5'-TTGTCCTAAGTTATGGAGCC-3'; Vpureverse: 5'-TTCTGCTCTTTCTATTAGTC-3'; Tat-reverse: 5'-CTGACTTCCTGGATGCTT-3'; Nef-reverse: 5'-TTTT GACCACTTGCCACCCA-3'; Rev-forward: 5'-GCTCATC AGAACAGTCAGAC-3'; Rev-reverse: 5'-CCTGACTCCA ATACTGTAGG-3' QRT-PCR results were normalized to GAPDH amplification levels and calculated by the 2-△△Ct comparative method.
-
HEK293 cells were seeded in a four-chamber slide 1 day prior to transfection with plasmid DNA expressing FLAGtagged CRTC2, CRTC2-S171A, or CRTC2-S171D. The cells were fixed with 4% paraformaldehyde (in 1xphosphate-buffered saline) for 10 min at room temperature and permeabilized with 0.1% Triton X-100 for 10 min at room temperature. The cells were then stained for 2 h at room temperature with antibodies against FLAG (1:500 dilution, Rabbit). After washing with 1Xphosphate-buffered saline, cells were incubated with Alexa Fluor 488-conjugated secondary anti-rabbit antibody (1:2000 dilution; Invitrogen). Nuclei were stained with DAPI (1 lg/mL in 1×phosphate-buffered saline). The images were recorded using the Zeiss Pascal laser scanning confocal microscope.
-
HEK293 cells were co-transfected with pNL4-3-luc-E- and CRTC2 plasmid DNA. Twenty hours after transfection, cells were treated with actinomycin D for 0, 0.5, 1, 2, 4, or 8 h. Total cellular RNA was extracted and analyzed by qRT-PCR using probes specific for HIV-1 gag and cellular GAPDH as described above.
-
Jurkat E4 and 2D10 cells were washed twice with 1 × PBS, and resuspended in Fixation/Permeabilization solution (B & D) for 20 min at 4 ℃. After washing twice in Perm/Wash buffer (B & D), cells were resuspended in Perm/ Wash buffer for 15 min at 4 ℃, then analyzed by flow cytometry on a FACS Calibur in the Flow Cytometry Core Lab in Cancer Hospital Chinese Academy of Medical Sciences. Data analysis was performed using FlowJo version 7.6. GFP-expressing cells were sorted using the defined gate, and the percentage of GFP-positive cells was determined.
-
Data are presented as mean ± standard deviation (SD) from at least three independent experiments unless otherwise indicated. Comparisons between groups were analyzed using the two-tailed Student's t test (two groups) or a one-way ANOVA with Dunnet's post hoc test (multiple groups). P values < 0.05 were considered statistic significant.
Cells, Viruses, Plasmids and Reagents
Western Blots
shRNA Knockdown
HIV-1 Infection Assay
Infection Assay of IAV, HCV and ZIKV
Luciferase Reporter Assay
ChIP Assay
Quantification of HIV-1 RNA and Integrated DNA
Immunofluorescence Microscopy
RNA Stability Assay
Flow Cytometry
Statistical Analysis
-
To investigate whether CRTC2 is involved in HIV-1 infection, we first examined the effect of HIV-1 infection on the expression of endogenous CRTC2, and found a significant increase (fourfold) of CRTC2 expression in PBMCs (Fig. 1A) and a moderate increase (0.5 fold) in HEK293T cells (Fig. 1B) upon the infection with VSVGpseudotyped HIV-1 (NL4-3-Luc), as shown by results of Western blots. To test the potential effect of CRTC2 on HIV-1 infection, we first transfected HEK293T cells with varying amount of plasmid expressing Flag-tagged CRTC2, then infected the transfected cells with the VSVGpseudotyped HIV-1 reporter virus NL4-3-Luc. The level of HIV-1 infection was determined by measuring luciferase activity in the infected HEK293T cells. HIV-1 infection was inhibited by up to approximate 70% by CRTC2 in a dose-dependent manner (Fig. 1C). This suggests that CRTC2 acts as an inhibitor of HIV-1 infection. In support of this result, HIV-1 infection increased by 1.5-fold in CRTC2-knockdown (KD) HEK293T cell line stably expressing CRTC2-specific short hairpin RNAs (shRNAs), compared with that in the control cell line expressing a scramble shRNA sequence (Fig. 1D). We further tested the effect of CRTC2 expression on several HIV-1 strains, including NL4-3, BH10 and AD8, by transfecting HEK293T cells with HIV proviral DNA constructs and CRTC2 DNA. After 48 h, levels of viruses in the cell culture supernatants were determined by infecting the TZM-bl indicator cells. The result showed that the overexpressed CRTC2 reduced virus production of different HIV-1 strains by about 45%–85% (Fig. 1E). Together, these data suggest that in response to HIV-1 infection, cells increase the expression of HIV-1 inhibitory protein CRTC2 as a cell protection mechanism.

Figure 1. CRTC2 inhibits HIV-1 infection. A, B PBMC and HEK293T cells were infected with VSVG-pseudotyped HIV-luc. 48 h later, levels of CRTC2 and HIV-1 Gag were determined by western blotting (upper panel). The relative CRTC2 and actin levels were quantified by ImageJ software (lower panel). C Effects of CRTC2 overexpression on HIV-1 infection. HEK293T cells were transfected with CRTC2 (0, 200, 500, 1000 ng), using pcDNA3.1 to keep constant the total amount of plasmids, and then were infected with VSVGpseudotyped HIV-luc. 48 h later, levels of luciferase activity in the infected cells were measured to evaluate HIV-1 infection. D Effects of CRTC2 knockdown on HIV-1 infection. CRTC2 was stably knocked down in HEK293T cells, then CRTC2-KD cells were infected with VSVG-pseudotyped HIV-luc. Levels of luciferase activity in the infected cells were measured to evaluate HIV-1 infection. E Effects of CRTC2 overexpression on the viral production of different HIV-1 strains. HEK293T cells were co-transfected with 100 ng HIV-1 cDNA and 400 ng CRTC2 plasmid DNA. 48 h later, levels of infectious HIV-1 thus produced were determined by infecting the TZM-bl indicator cells. F Effects of CRTC1, CRTC2 and CRTC3 on HIV-1 infection. HEK293T cells were transfected with 500 ng CRTC1, CRTC2 or CRTC3 plasmid DNA, then infected with VSVG-pseudotyped HIV-luc. After 48 h, levels of CRTC1, CRTC2 and CRTC3 in the cell lysates were determined by Western blots. pcDNA3.1 vector were used as control in the data. Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. The numbers below the gel represent the relative level of the indicated protein, which was determined by the intensity of each band normalized relative to its corresponding loading control band (actin) from three independent experiments using ImageJ. *denotes P < 0.05, **denotes P < 0.01, which were determined by the unpaired student t-test.
Since the members of the CRTC family share highly similar structures, we further tested whether CRTC1 and CRTC3 also inhibit HIV-1 infection. Indeed, HIV-1 infection was impaired to a similar extent in cells expressing either CRTC1, CRTC2 or CRTC3 (Fig. 1F), suggesting that inhibition of HIV-1 infection is a property shared by the CRTC family. Of note, although CRTC1 exhibited the strongest impact on HIV-1 infection, the predominant residence in the brain does not likely support its relevance. Moreover, CRTC2 and CRTC3 reduced HIV-1 infection to the similar level, the lower expression of CRTC2 suggests a stronger inhibition by CRTC2. We therefore selected CRTC2 in the following study.
-
We next investigated which step of HIV-1 life cycle was inhibited by CRTC2. First, we examined the effect of CRTC2 on HIV-1 DNA integration. To this end, CRTC2- KD or CRTC2-overexpressing HEK293T cells were infected with the VSVG-pseudotyped HIV-1, and the level of integrated HIV-1 DNA was determined by Alu-PCR as we described previously (Chun et al. 1997; Ma et al. 2018b). As shown in Fig. 2A, neither silencing (right panel) nor overexpressing (left panel) CRTC2 affected the level of the integrated viral DNA in the infected cells, compared with that of control groups. Thus, CRTC2 does not affect the early events of HIV-1 replication, including reverse transcription, nuclear import of viral DNA and viral DNA integration. We therefore further measured the level of viral gag RNA by qRT-PCR, and found that the overexpression of CRTC2 marked reduced viral gag RNA expression (Fig. 2B, left panel), which correlated with the decrease in Gag protein (Fig. 2C, left panel) and virus production (Fig. 2D, left panel). In agreement with the above observation, silencing CRTC2 increased the expression of viral gag RNA, as well as Gag protein and virus production (Fig. 2B–2D, right panels). These data suggest that CRTC2 inhibits HIV-1 RNA expression.
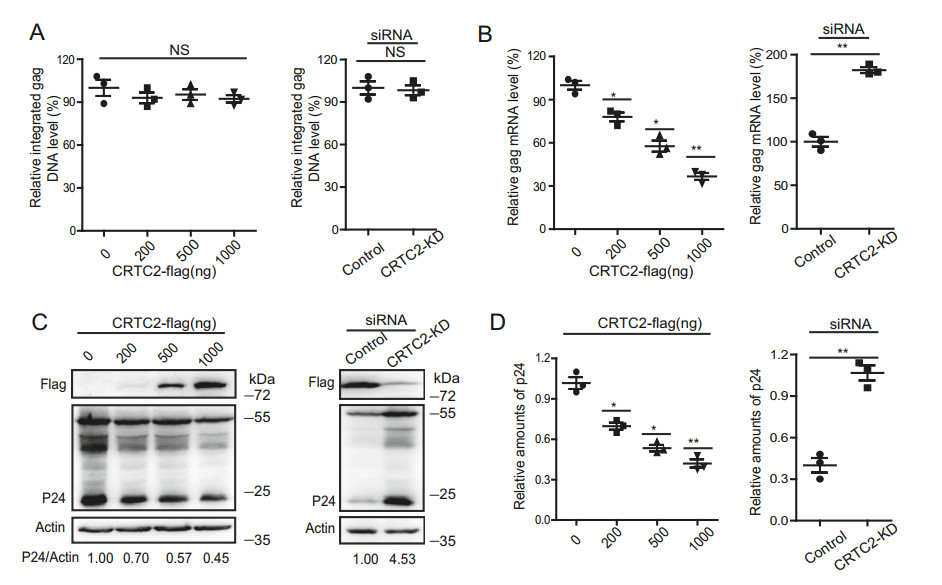
Figure 2. CRTC2 reduces HIV-1 RNA expression. A, B Effect of CRTC2 on HIV-1 DNA integration. VSVG-pseudotyped HIV-luc virus was used to infect HEK293T cells which either overexpressed increasing amounts of CRTC2 (200, 500, 1000 ng) (left panel) or had CRTC2 knocked down (right panel). Levels of integrated HIV-1 were determined with Alu-PCR (A), levels of HIV-1 gag mRNA were quantified with qRT-PCR (B). C, D Effects of CRTC2 on HIV-1 protein expression and HIV-1 production. HEK293T cells were cotransfected with CRTC2 DNA, pNL4-3-luc-E- and VSVG DNA. CRTC2-KD HEK293T cells were also transfected with pNL4-3-luc-Eand VSVG DNA. Expression levels of HIV-1 Gag/p24 and CRTC2 in the transfected cells were determined by western blotting, actin was used as a loading control (C). The numbers below the gel represent the relative level of p24, which was determined by the intensity of each band normalized relative to its corresponding loading control band (actin) from three independent experiments using ImageJ. Virus production in the culture supernatants from CRTC2 overexpressing cells (left panel) and CRTC2 KD cells (right panel) were measured with ELISA (D). Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. *denotes P < 0.05, **denotes P < 0.01, NS denotes not significant, which were determined by the unpaired student t-test.
-
In order to determine whether the CRTC2-mediated inhibition is specific to HIV-1, we assessed the effect of CRTC2 on the infection of influenza A virus (IAV), hepatitis C virus (HCV), Zika virus (ZIKV) and long interspersed nuclear elements 1 (LINE-1). CRTC2-overexpressing A549, Huh7.5.1, and Vero E6 cells were infected with IAV-1, HCV and ZIKV respectively, or transfected with the LINE-1 plasmid DNA. Levels of viral protein expression were then measured by Western blots using antibodies against IAV-1 NP, HCV Core, ZIKV NS3 or LINE-1 ORF1 protein. The results did not show any inhibition of the above viruses or LINE-1. In contrary, modest increase in the expression of ZIKV NS3 and HCV Core proteins was observed in response to CRTC2 overexpression (Fig. 3). These data demonstrate that CRTC2 exerts specific inhibition of HIV-1.
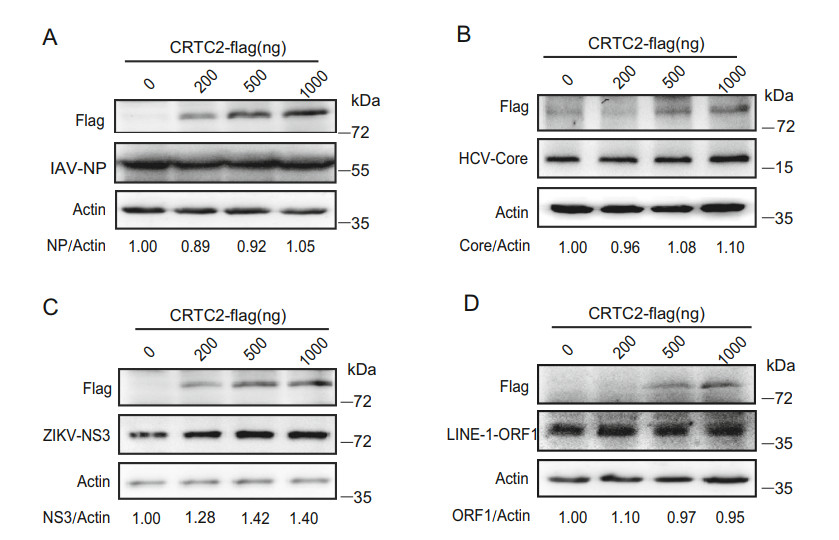
Figure 3. Effects of CRTC2 on the infection of IAV, HCV, and Zika virus. A549 cells, Huh7.5.1 cells and Vero E6 cells were transfected with 500 ng CRTC2 DNA, then infected with IAV-1 (A), HCV (B) or Zika virus (C), respectively. HEK293T cells was also co-transfected with CRTC2 and LINE-1 DNA (D). Levels of virus infections were assessed by measuring viral NP (A), Core (HCV) (B), NS3 (Zika virus) (C) and ORF1p (LINE-1) (D) in Western blots. One representative result from at least three independent experiments was shown. The numbers below the gel represent the relative level of the indicated protein, which was determined by the intensity of each band normalized relative to its corresponding loading control band (actin) using ImageJ.
-
CRTC2 contains four functional domains: N-terminal CBD, REG, SD and C-terminal TAD domains (Fig. 4A). We therefore generated CRTC2 mutants to determine which of these domains are involved in suppressing HIV-1 infection (Fig. 4A). HEK293T cells were co-transfected with the mutated CRTC2 DNA and HIV-luc/VSV-G. The levels of viral Gag protein, viral RNA in transfected cells and the production of progeny virus were determined. As shown in Fig. 4B–4D, deletion of either the N-terminal CBD or the C-terminal TAD did not markedly affect the anti-HIV-1 activity of CRTC2, suggesting the essential role of the other two domains, REG and SD. Of note, CBD of CRTC2 mediates its association with CREB and subsequently stimulates the expression of downstream genes after CRTC2 is translocated into the nucleus, the observation that CBD was not required for the anti-HIV-1 activity of CRTC2 suggest it functions independent of binding with CREB at least and likely via transcriptional trans-activation. Although the expression of 1-614aa mutant is usually much lower than wild type or other mutated CRTC2, we detected a reproducible decrease of Gag mRNA and viral production level. The expression of p55 level decreased mildly compared with the level of mRNA and viral production, probably because western blot is a semi-quantitative method and we consider mRNA and viral production level is a more reliable indicator for the anti-HIV activity of the mutant.
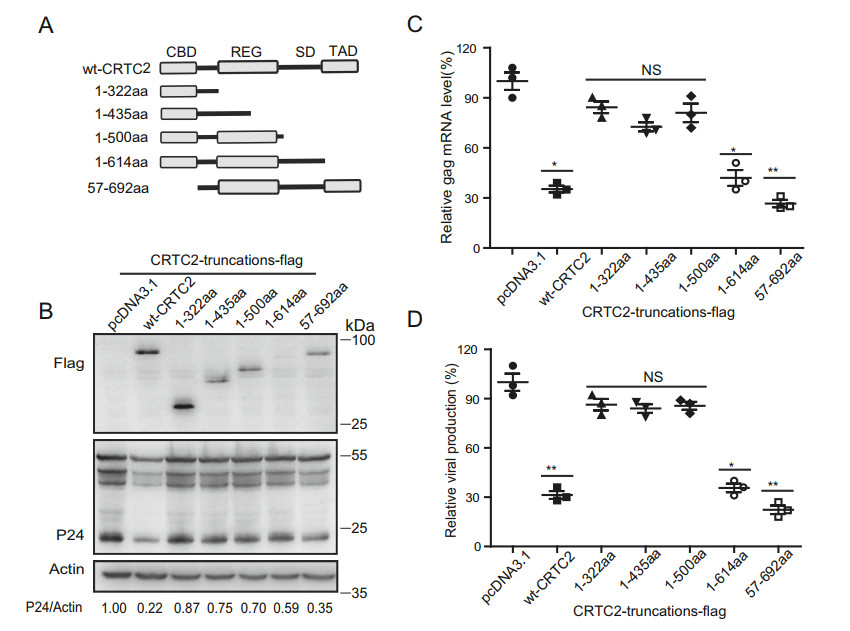
Figure 4. Motifs of CRTC2 essential for HIV-1 inhibition. A Illustration of CRTC2 truncations. B Effect of CRTC2 mutants on HIV-1 Gag/p24 expression. HEK293T cells were co-transfected with CRTC2 DNA, pNL4-3-luc-E- and VSVG DNA. Expression of HIV-1 Gag/p24 was determined by western blotting. The numbers below the gel represent the mean ± SD of band density of p24/actin from three independent experiments quantified by ImageJ. C Effect of CRTC2 mutants on HIV-1 RNA expression. D Effect of CRTC2 mutants on HIV-1 production. The nascent HIV-1 Luc reporter viruses in the supernatant were used to infect SupT1 cells. The viral production was determined by measuring luciferase activity. Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. *denotes P < 0.05, **denotes P < 0.01, NS denotes not significant, which were determined by the unpaired student t-test.
Without activation, CRTC2 is sequestered in the cytoplasm as a result of phosphorylation at Ser171 and Ser275 (Conkright et al. 2003). Dephosphorylation at these two sites, particularly Ser171, leads to CRTC2 nuclear translocation and activation of gene transcription (Altarejos and Montminy 2011). Indeed, we found that the Ser171Ala mutation (phosphorylation-defective) caused a predominant localization of CRTC2 in the nucleus, whereas the Ser171Glu mutant (phosphorylation-like) primarily resided in the cytoplasm (Fig. 5A). Upon the expression of these two mutants in HIV-1 infected cells (Fig. 5B), the Ser171Ala mutant showed strong inhibition of HIV-1 protein expression (Fig. 5B), viral RNA expression (Fig. 5C) and viral production (Fig. 5D), whereas the Ser171Glu mutant lost inhibition of HIV-1 infection (Fig. 5B–5D). These data suggest that the antiviral activity of CRTC2 depends on its localization to the nucleus, although we cannot rule out the possibility that dephosphorylation of Ser171 may affect functions of CRTC2 other than nuclear translocation.
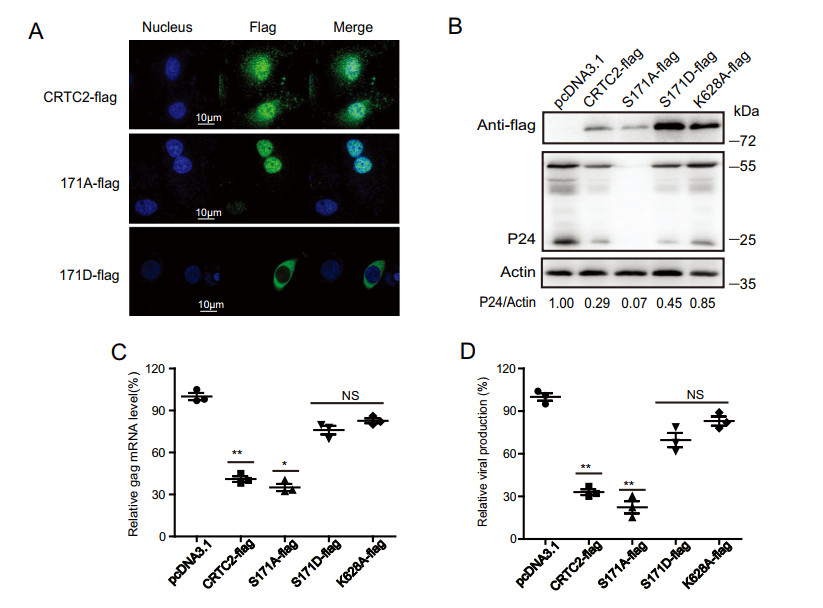
Figure 5. Effect of CRTC2 phosphorylation and ubiquitination on its inhibition of HIV-1 infection. A Effect of the S171 mutations on subcellular location of CRTC2. The S171A mutation abrogates serine phosphorylation, whereas S171D mimics constitutive phosphorylation at this site. Immunostaining images of wt CRTC2, S171A, and S171D mutants (green) were shown. B–D Effects of CRTC2 mutants on HIV-1 Gag/p24 expression (B), HIV-1 RNA expression (C) and HIV-1 production (D). HEK293T cells were co-transfected with CRTC2 DNA, pNL4-3-luc-E- and VSVG DNA. Levels of Gag/p24 were determined by western blotting, HIV-1 RNA expression was examined by qRT-PCR. Levels of HIV-1 luc reporter virus in the supernatant were determined by infecting SupT1 cells, followed by measuring luciferase activity. Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. *denotes P < 0.05, **denotes P < 0.01, NS denotes not significant, which were determined by the unpaired student t-test, the scale bar represents 10 μm.
In the context of inhibiting HIV-1 infection, we further examined Lys628 which has been reported to undergo ubiquitination and regulate CRTC2 activity (Liu et al. 2008). Our data showed that the K628A mutant did not affect HIV-1 gene expression, nor viral production (Fig. 5B–5D), suggesting that the anti-HIV-1 activity of CRTC2 is independent of Lys628 ubiquitination.
-
CRTC2 has been reported to modulate alternative RNA splicing through its SD motif (Bonni et al. 1995; Amelio et al. 2009). Given that the SD domain-deleted CRTC2 lost inhibition of HIV-1, we decided to investigate whether CRTC2 also interferes with HIV-1 RNA splicing, which may result in reduction in specific viral RNA species. To this end, we quantified the level of different viral RNA species in HIV-1 infected cells expressing either CRTC2 or the inactive mutant CRTC2.1–132. The results showed similar reductions in all viral transcripts examined, including unspliced mRNA (gag), single spliced mRNA (vif, vpu, vpr) and full spliced mRNA (rev, nef, tat), in the presence of CRTC2, whereas the CRTC2 mutant did not affect the expression of any viral RNA (Fig. 6A). These data demonstrate that CRTC2 diminishes the expression of all viral RNA rather than affects viral RNA splicing.
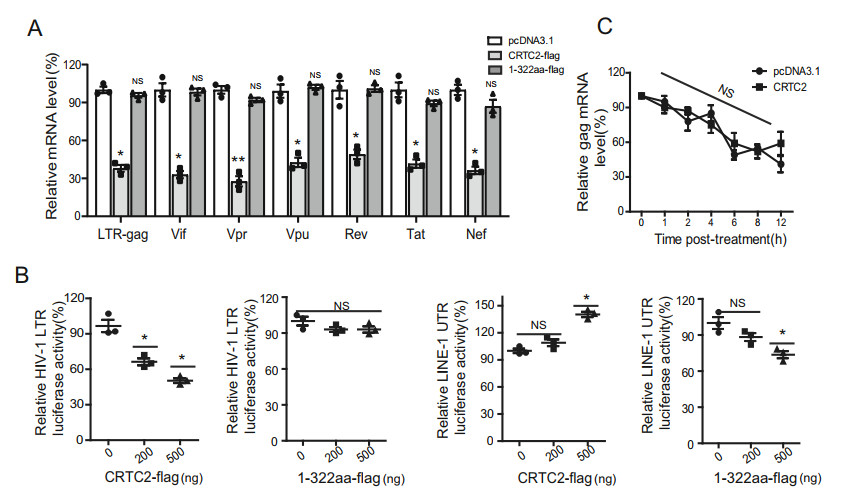
Figure 6. Effect of CRTC2 on HIV-1 RNA expression. A CRTC2 does not affect HIV-1 RNA splicing. HEK293T cells were co-transfected with, pNL4-3-luc-E-, VSVG DNA and wt CRTC2 or CRTC2.1-322 mutant. After 48 h, quantitative RT-PCR was performed to measure the levels of gag, vif, vpu, vpr, rev, nef and tat mRNA. B Effect of CRTC2 on the promoter activity of HIV-1 LTR. HEK293T cells were co-transfected with CRTC2 DNA and LTR-luc or LINE-1 50UTR-luc reporter DNA. CRTC2.1-322 mutant was also tested as a control. After 48 h, Levels of luciferase activity in the transfected cells were measured. C Effect of CRTC2 on the stability of HIV-1 mRNA. HEK293T cells were co-transfected with pNL4-3-luc-E- and CRTC2 DNA. Twenty hours after transfection, cells were treated with actinomycin D for 0, 0.5, 1, 2, 4, or 8 h. Levels of HIV-1 RNA was determined by qRT-PCR. Transfection with pcDNA3.1 vector DNA serves as the control. Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. *denotes P < 0.05, **denotes P < 0.01, NS denotes not significant, which were determined by the unpaired student t-test.
The overall reduction of HIV-1 RNA expression may result from impairment of transcription from HIV-1 LTR promoter. We therefore examined the effect of CRTC2 on HIV-1 LTR transcription using a luciferase reporter DNA construct in which expression of the luciferase gene is under the control of the HIV-1 LTR promoter (Koseki et al. 1998). Upon expression of CRTC2, we observed a marked reduction of the transcription of HIV-1 LTR (Fig. 6B, first panel from left), but an increase in LINE-1 5'-UTR promoter transcriptional activity (Fig. 6B, third panel from left). In contrast, the CRTC2.1–132 mutant, which does not inhibit HIV-1 infection (Fig. 4D), showed no effect on luciferase expression from HIV-1 LTR (Fig. 6B, second panel from left). These data suggest a selective inhibition of HIV-1 promoter by CRTC2.
To rule out the possibility that CRTC2 induces degradation of HIV-1 RNA, we measured the decay kinetics of viral RNA in HIV-infected cells using actinomycin D to suppress RNA Pol Ⅱ (Li et al. 2013). Similar decay kinetics of HIV-1 RNA were observed with or without CRTC2 expression, suggesting that CRTC2 does not affect the stability of HIV-1 RNA (Fig. 6C).
-
HIV-1 transcription starts with RNA Pol Ⅱ binding to LTR. But RNA Pol Ⅱ pauses immediately downstream the transcription initiation site, until P-TEFb is recruited by Tat and hyperphosphorylates the CTD of RNA Pol Ⅱ, which releases the paused RNA Pol Ⅱ and allows productive transcription elongation (Karn and Stoltzfus 2012). To understand how CRTC2 inhibits HIV-1 transcription, we performed qRT-PCR to measure the amount of transcripts that were proximal and distal to the LTR, which reflects the efficiency of HIV-1 transcription initiation and elongation, respectively. The results showed that both HIV-1 transcription initiation and elongation were inhibited to similar extent by CRTC2 (Fig. 7A). One interpretation of this observation is that CRTC2 inhibition of transcription initiation mainly accounts for the reduction in HIV-1 RNA expression.
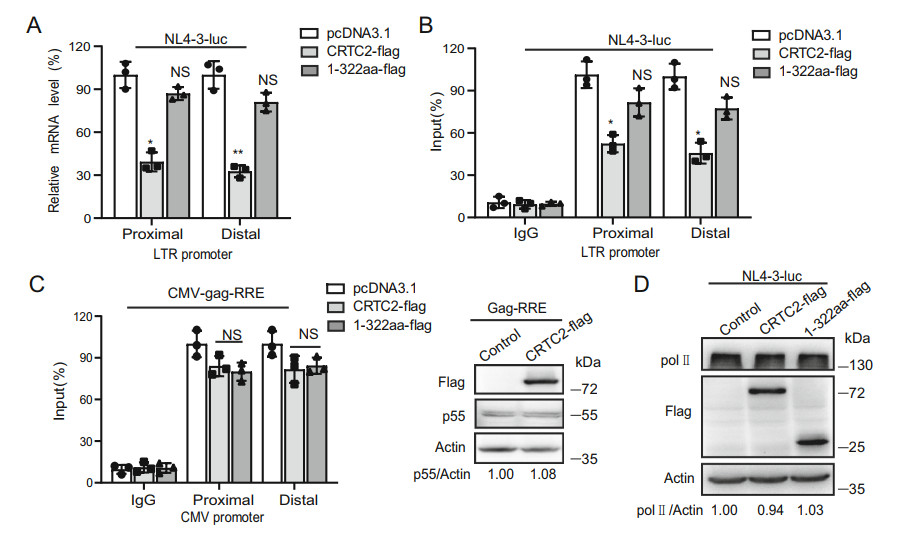
Figure 7. CRTC2 prevents RNA Pol Ⅱ from binding to HIV-1 LTR. A HEK293T cells were co-transfected with pNL4-3-luc-E- and CRTC2 or CRTC2.1-322 mutant. After 48 h, quantitative PCR was performed to measure the amount of HIV-1 RNA using primers that amplify RNA sequences either proximal or distal to the transcription start site. B CHIP-qPCR was performed to determine the levels of RNA Pol Ⅱ that is associated with HIV-1 genomic regions either proximal or distal to the transcription start site. C HEK293T cells were co-transfected with CRTC2 and CMV-gag-RRE DNA. Levels of RNA Pol Ⅱ at the CMV promoter was determined by (ChIP)-qPCR. Gag protein expression was analyzed by western blots. D HEK293T cells were co-transfected with CRTC2 and pNL4-3-luc-E- DNA. The protein expression was analyzed by western blots. Data were presented as mean ± standard deviations (SD). One representative result from at least three independent experiments was shown. *denotes P < 0.05, **denotes P < 0.01, NS denotes not significant, which were determined by the unpaired student t-test.
Next, we investigated whether CRTC2 affects RNA Pol Ⅱ occupancy at HIV-1 promoter by performing chromatin immuno-precipitation (ChIP)-qPCR using anti-RNA Pol Ⅱ antibodies. The results showed that CRTC2 decreased RNA Pol Ⅱ occupancy at both the proximal and distal regions of HIV-1 genome by more than 50% (Fig. 7B). In contrast, the CRTC2.1–322 mutant marginally affected RNA Pol Ⅱ occupancy at HIV-1 LTR. As a control, CRTC2 did not affect RNA Pol Ⅱ occupancy at the CMV promoter, and as a result, did not affect the expression of Gag protein from this promoter (Fig. 7C) (Galvin et al. 2000; Suzuki et al. 2018). Furthermore, CRTC2 did not affect the total amount of RNA Pol Ⅱ (Fig. 7D), which rules out the possibility that the diminished RNA Pol Ⅱ occupancy at HIV-1 LTR is a result of reduced RNA Pol Ⅱ expression. Together, these data indicate that CRTC2 inhibits transcription from HIV-1 LTR by diminishing RNA Pol Ⅱ occupancy.
-
HIV-1 latency is characterized mainly by silencing of transcription from a provirus (Lassen et al. 2004; Coiras et al. 2009; Margolis, 2010; Trono et al. 2010). The inhibitory effect of CRTC2 on HIV-1 transcription suggests a potential role for CRTC2 in HIV-1 latency. We therefore investigated the effect of CRTC2 on the activation of an integrated HIV-1 DNA in a T cell clone Jurkat E4, a CD4+ T cell line harboring an HIV-1 proviral DNA derived from HIV-1NL4-3, which expresses the wild-type Tat protein and carries the fluorescent d2EGFP protein in place of the Nef gene as a marker of transcriptional activation (Pearson et al. 2008). The provirus is integrated into the fourth exon of the centromere protein P gene (CENPP) on chromosome 9. Jurkat E4 cells express GFP when HIV-1 is activated from latency by tumor necrosis factor α (TNF-α), Vorinostat (SAHA) or trichostatin A (JQ-1). We observed that when the endogenous CRTC2 was knocked down with shRNA (Fig. 8A), GFP expression increased by approximate 44% in the absence of any stimulation (Fig. 8B), suggesting a possible role of CRTC2 in maintaining HIV-1 latency. In support of this finding, overexpression of CRTC2 suppressed JQ-1- mediated activation of latent HIV-1 by more than 30% (Fig. 8C, 8D). The same experiment was carried out with 2D10 cell line (Pearson et al. 2008) that was widely used as HIV-1 latency model, and a similar change was observed (data were not shown). These results suggest a role of CRTC2 in maintaining HIV-1 latency through suppression of HIV-1 gene expression.
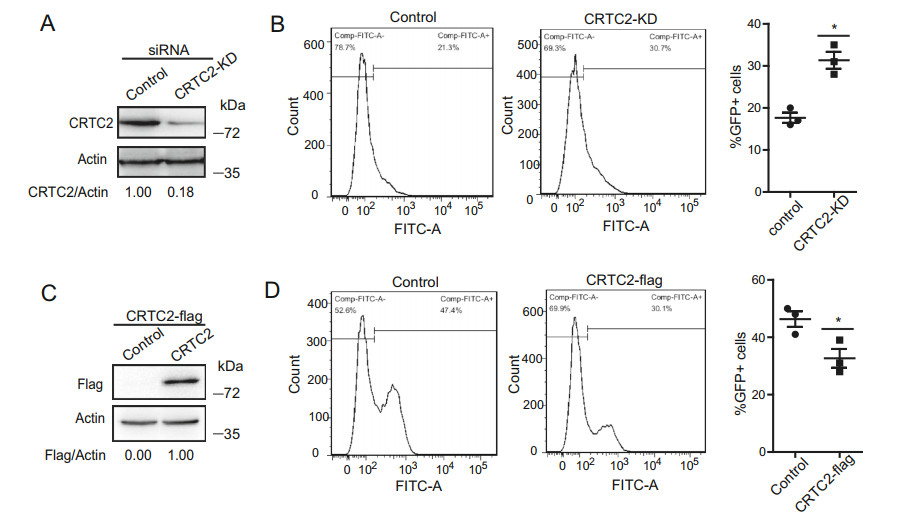
Figure 8. CRTC2 suppresses activation of latent HIV-1 DNA. A Jurkat E4 cells were electro-transfected with CRTC2-specific shRNA or a negative control. Knockdown of CRTC2 was confirmed by western blotting. B Percentage of GFP-positive CRTC2-KD cells (middle panel) or control cells (left panel) were determined by flow cytometry. The ratio of GFP+ cells were quantified (right panel). C Jurkat E4 cells were electro-transfected with CRTC2-flag DNA or control vector. Expression of CRTC2 was examined by Western blotting. D CRTC2-overexpression Jurkat E4 cells were treated with JQ1 for 24 h. GFP-positive control cells (left panel) and CRTC2-KD cells (middle panel) were scored by flow cytometry. The ratio of GFP+ cells were quantified (right panel). Results shown represent mean ± SD of three independent experiments, *denotes P < 0.05.
CRTC2 Suppresses HIV-1 Infection
CRTC2 Inhibits HIV-1 RNA Expression
CRTC2 Does Not Affect the Infection of IAV, HCV and ZIKV
Key Motif and Amino Acid Essential for CRTC2 to Inhibit HIV-1
CRTC2 Inhibits the Promoter Activity of HIV-1 LTR
CRTC2 Prevents RNA Pol Ⅱ from Binding to HIV-1 LTR
CRTC2 Suppresses Activation of Latent Proviral DNA
-
CRTCs are transcription activators, enhancing the expression of CREB-targeted genes through binding to the bZIP domain of CREB protein (Bonni et al. 1995; Galvin et al. 2000; Jiang et al. 2009). In addition to regulating cellular gene expression in response to cAMP and other signals, CRTCs also affect the replication of several viruses. For example, CRTC2 enhances HBV transcription and replication by inducing PGC1a expression (Conkright et al. 2003). It also activates Epstein-Barr virus from latency through interaction with viral BZLF1 protein (Murata et al. 2009). In contrast to these stimulatory effects on virus infection, our study showed that CRTC2 inhibited HIV-1 transcription and production, which likely contributes to the maintenance of HIV-1 latency. This inhibitory effect on HIV-1 appears to be specific, since we tested several RNA viruses and retrotransposon LINE-1, and none of them was affected by CRTC2. It will be interesting to test whether CRTC2 also suppress transcription from the LTR of endogenous retroviruses, which may reveal a role of CRTC2 in silencing the retroviral DNA in our genome.
Host factors play key roles in controlling HIV-1 gene expression and determine the transition between viral latency and active replication. Transcriptional factors, NF-κB, NFAT (nuclear factor of activated T cells), and C/EBPβ, bind to the LTR promoter and stimulate transcription (Rafati et al. 2011; Van Duyne et al. 2011; Schiralli Lester and Henderson 2012). Not surprisingly, some factors were reported to inhibit HIV-1 transcription, including CBF1 (C-promoter binding factor-1) (Tyagi and Karn 2007), NELFCD (negative elongation factor) (Zhang et al. 2007), and HDACs (histone deacetylases) (Keedy et al. 2009), although their function in transcription suppression is relatively general, not specific to HIV-1. Our work demonstrates that CRTC2 functions as a specific inhibitor of HIV-1 transcription, through reducing RNA Pol Ⅱ occupancy at LTR. In light that host proteins Naf1 (Nef-associated factor 1) (Li et al. 2016), SAFB1 (scaffold attachment factor B) (Ma et al. 2018a), SUN2 (SAD1/UNC84 domain protein-2) (Sun et al. 2018) and SMYD2 (SET and MYND domain-containing protein 2) (Boehm et al. 2017) inhibit HIV-1 gene expression through different mechanisms, it appears that cells exert multiple layers of control on HIV-1 transcription. We noted that HIV-1 infection increased CRTC2 expression, especially in PBMCs (Fig. 1A, 1B). It is reasonable to propose that CRTC2 represents one cellular protection mechanism that responds to and inhibits HIV-1 infection. However, by suppressing HIV-1 gene expression, CRTC2 contributes to driving HIV-1 into latency. This latter possibility awaits further investigation of CRTC2 level in HIV-1 latent cells.
Our mechanistic study showed that CRTC2 reduced RNA Pol Ⅱ occupancy at the HIV-1 promoter (Fig. 7), which represents a new mechanism of CRTC2 action. CRTC2 has been reported to regulate the transcription and splicing of CREB target genes in a CREB-dependent fashion (Amelio et al. 2009). In contrast, we observed that CRTC2 still inhibited HIV-1 when its CBD domain was deleted (Fig. 4C, 4D). Since CRTC2 interacts with CREB via its CBD domain, it is concluded that CREB is not required for CRTC2 to inhibit HIV-1. In support of this notion, CRTC2 inhibition of HIV-1 is independent of ubiquitination at Lys628 (Fig. 5B–5D), a post-translational modification which has been shown essential for CRTC2 activation of CREB target genes. Nevertheless, we can not rule out the possibility that CRTC2 binding to the LTR can also block upstream steps in the transcription cycle such as pre-initiation complex assembly. Moreover, we observed that CRTC2 reduced the levels of all HIV-1 transcripts to the same extent (Fig. 6A), thus ruling out the effect on HIV-1 RNA splicing. The detailed mechanisms how CRTC2 reduce RNA Pol Ⅱ occupancy awaits further investigation.
Interestingly, in contrast to inhibition of HIV-1, previous work showed that CRTC2 enhances transcription of HTLV-1, a close member of HIV-1 (Rabbi et al. 1997). CRTC1 and CRTC2 together promote Tax activation of HTLV-1 LTR (Rafati et al. 2011). The Tax protein cooperates with phosphorylated CREB, CRTC2 and p300 to activate CRE-dependent Cyclin D1 transcription (Jiang et al. 2009). HIV-1 Tat protein recruits p300/CBP and PCAF to viral LTR, which is essential for HIV-1 gene expression when the provirus integrates in transcriptionally inactive regions of the genome (Tian et al. 2014). Despite the similarity in transcriptional regulation between these two viruses, a recent work reported that TPA activated transcription o fHTLV-1 LTR through recruiting Sp1, p53 and CREB, whereas TPA activation of HIV-1 LTR depended on the PKC activity via the NF-κB pathway (Jabareen et al. 2018). The different requirement of cellular transcription factors for transcription of HIV-1 and HTLV-1 may explain the opposing effects of CRTC2 on these two viruses.
In summary, we have identified a novel role of CRTC2 in suppressing HIV-1 transcription and replication. These results enrich our understanding of cellular regulation of HIV-1 transcription and latency, and suggest new avenues for the development of HIV cure strategies.
-
We thank National Infrastructure of Microbial Resources (NIMR-2014-3) for providing valuable reagents. This work was supported by the National Mega-Project for Infectious Disease (2018ZX10301408 SC), the National Key Research and Development program of China (2018YFE0107600 SC), the National Natural Science Foundation of China (81903679 LM), the National Natural Science Foundation of China(81772205 SC), Peking Union Medical College Youth Fund (332017075 LM), CAMS innovation fund for Medical Sciences (2018-I2M-3-004 SC).
-
SC, JD and LM designed the study and wrote the paper. LM, SC and ZW performed the major part of the experiments. SG, JZ, DY, QL, ZL, XL, FG and CL contributed to data analysis. SC and JD finalized the manuscript. All authors read and approved the final version of the manuscript.
Author Contributions
-
The authors declare that they have no conflict of interests.
-
This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: