











-
The Human immunodeficiency virus type-1 (HIV-1) Vif protein is essential for virus replication in non-permissive cells such as macrophages, primary human T-cells, and some restrictive T-cell lines (8, 32). Non-permissive cells express APOBEC3G, a cellular protein that actively blocks retroviral infection but which is counteracted by HIV-1 Vif (37, 42).
APOBEC3G is a member of the cellular cytidine deaminase family, which has been implicated in the restriction of a broad range of exogenous retroviruses (9, 10, 12), endogenous retroviruses (5, 6), and the hepadnavirus hepatitis B (35). Vif counteracts APOB-EC3G by inhibiting its translation or intracellular half-life (14, 34) and inducing its polyubiquitination and proteasomal degradation by a specific interaction with APOBEC3G. The deaminase is recruited to degradation as part of the Vif-Cul5-SCF complex through a novel SOCS (suppressor of cytokine signaling)-box and an HCCH motif present in Vif, essential for the recruitment of Cul5 to the complex (4, 18, 21, 24, 25, 38, 39). The increased degradation and/or reduced level of APOBEC3G expression by Vif excludes APOBEC3G from incorporation into virions (14, 17)and its consequent absence during reverse transcription in the target cell allows the virus to replicate.
We wanted to develop a new simple cellular system for studying the functions of Vif, APOBEC3G and their interactions. So in this study, the subcellular localization of Vif, APOBEC3G were observed in S. pombe cells. To evaluate if the HIV-1 Vif protein can specifically recognizes and interfere with APOBEC3G in fission yeast cells, Vif protein was over-expressed by using codon optimization, and the APOBEC3G protein was co-expressed with Vif protein in the same S. pombe cell. Our results showed that although Vif and APOBEC3G evolved in a different cellular environment, Vif mediated the degradation of APOBEC3G in S. pombe cells; thus these results support the proposal that S. pombe cells are suitable model cells for the study of Vif, APOBEC3G and their interactions.
HTML
-
A wild-type S. pombe strain, SP223 (h- ade-216 leu1-32 ura4-294), was regularly maintained in YEA media (0.5% Difco yeast extra, 3% glucose, and adenine [225μg/mL)). Selection media for growth of S. pombe cells containing the expression vectors or recombinant constructs included standard Edinburgh minimal medium(EMM) supplemented with uracil, adenine, and leucine, all at 225μg/mL, as needed. Thiamine, prepared in distilled water as a 20mmol/L stock solution of thiamine hydrochloride, was added to the medium at a final concentration of 20μmol/L. All liquid cultures were grown at 30℃, with constant shaking at 200 rin/m. Agar plates were incubated at 30℃ for 3 to 4 days to obtain individual colonies.
-
Plasmids pYZ1N, pYZ3N and pYZ2N were generous gifts from Dr. Richard Yuqi Zhao, which have been described previously (41) and were used to express proteins in fission yeast. They were derived from pREP series vectors, which are widely used with fission yeast to express protein (22, 23). The gene corresponding to the protein of interest is cloned after the thiamine repressible nmt1 (no message in thiamine) promoter. The gene expression can be repressed or induced in the presence or absence of thiamine, respectively (1, 22). pYZ1N and pYZ3N carry the gene leu2 as a selection marker. Yeast transformed with those plasmids can grow in medium without the amino acid leucine. The difference between pYZ1N and pYZ3N plasmids is that pYZ1N allows the expression of the inserted gene alone and pYZ3N allows the expression of the inserted gene in fusion with GFP gene. pYZ2N is the same as pYZ1N except that there is a ura4 gene instead of leu2 gene to provide for the growth of yeast without uracil.
-
The vif gene of the HIV-1 viral isolate CNHN24 was amplified from reverse transcripted viral genome RNA using oligonucleotides 5' > gcgcgcgcgcccgcggat ctcgagatggaaaacagatggcagg > 3', 5' > gcgcgcgcgcgtcgactagtgtccactcattgt > 3'. The APOBEC3G gene was amplified with reverse transcripted H9's mRNA as templates using oligonucleotides 5' > gcgcgcgcgcccgcggatctcgagatgaagcctcacttcag > 3', 5' > gcgcgcgcgcgtcgactagttttcctgattctgg > 3'. The vif gene and APOBEC3G genes were cloned into fission yeast expression vectors separately at the sites labeled in those primers. The vif-s gene, in which the vif gene was optimized according to the codon used in yeast cells, was synthesized by the Shanghai Boya company. Insertion of the genes into vectors was confirmed by extracting DNA from individual Escherichia coli colonies containing the plasmid and subsequently analyzing the DNA by restriction mapping and PCR. The complete nucleotide sequence of the vif gene and APOBEC3G gene was confirmed by DNA sequencing. One clone was chosen and designated pYZ3N-vif, plasmid expresses Vif in fusion with GFP, pYZ3N-APOB-EC3G, plasmid expresses APOBEC3G in fusion with GFP, pYZ2N-vif, plasmid expresses Vif, pYZ3N-vif-s, plasmid containing optimalized vif gene for ex-pressing Vif in fusion with GFP, pYZ2N-vif-s and plasmid containing optimalized vif gene for expres-sing Vif.
-
Transformation of S. pombe was carried out by the lithium acetate procedure as described elsewhere (27). Briefly, sp223 cells were grown in EMM media with the supplements of adenine, leucine or uracil according to the selection requirements of each particular strain. For selection of pYZ1N or pYZ3N, cell cultures were grown in selective medium containing 225mg/L adenine and 225mg/L leucine. For selection of pYZ2N derived constructs, cell cultures were grown in selective medium containing 225mg/L adenine and uracil. For selection of strains containing both pYZ3N and pYZ2N plasmids, cell cultures were grown in selective medium containing 225mg/L adenine.
-
Cells containing plasmids were grown to stationary phase in 5 mL of EMM containing thiamine, with constant shaking at 30℃. A 1-mL aliquot of culture was collected, washed three times with distilled water to remove thiamine, and reinoculated into 5 mL of EMM medium at a concentration of 2×105 per mL cells with or without thiamine. Cells were collected at set time points. Subcellular localization of proteins in fusion with GFP were visualized with fluorescent microscopy, expressed proteins were harvested with detergent (PIERCE company) according to the protocol supplied by the company.
-
Nucleuses of S. pombe cells were visualized using DAPI (4', 6-diamidino-2-phenylindole) staining. In briefly, cell cultures were spun down in a microfuge at 3 000 r/min for 5min. Cellular sediment was resu-spended in EMM medium, 5µL cellular suspension was pipetted onto a slide and heated at 70℃ by putting the slide on a hot plate for 1min to fix the cells. The slide was then cooled for few seconds before adding 5µL 1× DAPI stock (DAPI 1µg/mL, p-pheny-lenediamine 1mg/mL, glycerol 90%), covered with a coverslip and viewed as a blue color under fluorescence microscopy.
-
For Western blot analysis, fission yeast cells were harvested and lysed in detergent (PIERCE company) and supplied with complete protease inhibitors (Roche Applied Science). Cellular lysates were prepared and the protein concentration was determined using the Pierce protein assay kit. For immunoblotting, an aliquot of total lysate (50 µg of proteins) in 5 ×SDS-PAGE sample buffer (4:1 v/v) was electrophoresed and transferred to a nitrocellulose filter. Filters were incubated with appropriate primary antibody in Tris-buffered saline (TBS, pH 7.5) and 5% skim milk in 4℃ overnight. After washing, the filter was further incubated with secondary antibody in TBS-Tween-20 (TBS-T) buffer for 1 h. Protein bands were visualized using ECL Western Blotting Detection Reagents. Goat anti-mouse or anti-rabbit IgG-HRP conjugate (dilution of 1:3 000) were used as secondary antibodies according to the corresponding primary antibodies.
Yeast and media
Vectors for S. pombe expression
Molecular cloning of the vif, vif-s and APOBEC3G gene
Transformation of fission yeast
Expression of proteins in fission yeast
Localization of nucleus by DAPI staining
Western Blot Analysis
-
To examine the subcellular localization of HIV-1 Vif protein in fission yeast, pYZ3N-vif plasmid was transformed into sp223 with the lithium acetate method. GFP-Vif fusion protein was expressed by eliminating the thiamine in the media. Yeast cells were harvested 24 h after induction of protein's expression. Protein localization was visualized directly in live cells. Vif was localized predominantly to the nucleus in most of the cells (Fig. 1 A).

Figure 1. Subcellular localization of Vif and APOBEC3G proteins in S. pombe cells. Strains containing pYZ3N, pYZ3N-vif or pYZ3N-APOBEC3G were cultured in EMM selective media under the conditions of with or without thiamine. Those strains for induced for 24 h and live cells were harvested and resuspended in proper volume water and examined using fluorescence microscopy to observe localization of the GFP-tagged proteins, DNA was detected by staining with DAPI after cells were fixed as described in materials and methods. The scale bar represents 10µm. A: S. pombe cells containing pYZ3N-vif plasmid or pYZ3N were visualized under fluorescent microscopy. GFP panel: cells containing pYZ3N plasmid; Vif panel: Cells containing pYZ3N-vifplasmid. B: S. pombe cells containing pYZ3N-APOBEC3G plasmid or pYZ3N were visualized under fluorescent microscopy. GFP panel: cells containing pYZ3N plasmid; APOBEC3G panel: Cells containing pYZ3N-APOBEC3G plasmid.
To examine the localization of the APOBEC3G protein in fission yeast, APOBEC3G was expressed in fusion with GFP. 24 h after induction of protein's expression, yeast cells were harvested and visualized directly using fluorescent microscopy. APOBEC3G was localized throughout the cytoplasm and was largelyexcluded from the nucleus (Fig. 1 B). Most of the APOBEC3G was concentratedat the punctate bodies that were often in close proximity to thenucleus but were not necessarily restricted from other regionsin the cytoplasm (Fig. 1 B).
-
In previous studies, we found that the level of Vif protein expressed in fission yeast is very poor, so we tried using codon optimalization to increase the expression level of Vif protein in fission yeast. Gene encoding Vif was optimized for yeast codon usage (33) and synthesized by the Shanghai Boya company. After optimization, the G+C was decreased from 42.5% to 32.5%, which is closer to the observed value for yeast genes. The overexpression vector, pYZ3N-vif-s, was constructed successfully by the procedures described in Materials and methods. The recombinant plasmid was verified by PCR with Taq DNA polymerase, restriction endonuclease cleavage (data not shown) and sequenced. S. pombe cells which can over-express GFP-Vif protein was constructed by transforming fission yeast strain Sp223 with plasmidpYZ3N-vif-s. Proteins were induced for expression as described in Materials and Methods. Since Vif proteins were expressed in fusion with GFP, the level of Vif protein in single S. pombe cells can be estimated from the intensity of green fluorescence. In the lower panel of Fig. 2 A, the intensity of green fluorescent dots in the cells was markedly stronger than those in the top panel. Thus, through codonoptimization, GFP-Vif expression was strongly enhanced. In addition to the observation of fluorescence, immunoblot analysis was used to compare the concentration of GFP-Vif proteins in different cells, proteins expression was induced for 48 hours before the cells were harvested and lysed. Proteins were analyzed using Western Blot analysis after quantifi-cation with the PIERCE protein assay kit. ~50-kDa protein bands were detected with anti-GFP mono-clones in Vif-induced cells (Fig. 2 B, lane 2 and4). No specific bands were detected in either GFP-Vif-repressed cells (Fig. 2 B, lane 1 and 3).
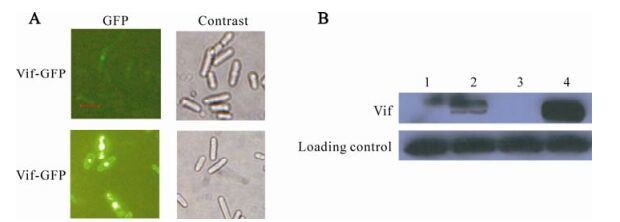
Figure 2. Codon optimization of vif gene significantly enhances the expression of Vif proteins in S. pombe cells. A: Strains containing pYZ3N-vif or pYZ3N-vif-s plasmids were induced to express protein for 24 h by diminishing the thiamine in media (-T) before being observed under fluorescent microscopy. Vif-GFP: S. pombe cells containing pYZ3N-vif plasmid; Vif-s-GFP: S. pombe cells containing pYZ3N-vif-s plasmid. The scale bar represents 10µm. B: Strains containing pYZ3N-vif or pYZ3N-vif-s were induced for protein expression for 48 h before cells were harvested and lysed to prepare cellular proteins. Equal quantities of proteins were subjected to electrophoresis and immunoblotting analysis. Vif protein levels were detected with anti-GFP antibody. Levels of α-Tubulin were used as protein loading controls. Lane 1, pYZ3N-vif/SP223 +T; Lane 2, pYZ3N-vif/SP223–T; Lane 3, pYZ3N-vif-s/SP223 +T; Lane 4, pYZ3N-vif-s/SP223–T.
-
One of Vif's main functions is mediating the degradation of APOBEC3G. So the ability of HIV-1 Vif to mediate the degradation of APOBEC3G in fission yeast was assessed by fluorescence observation and Western Blot assay. To do this, GFP-APOBEC3G fusion protein expressing plasmid pYZ3N-APOBEC3G was transformed together with Vif protein expressing plasmid pYZ2N-vif, pYZ2N-vif-s or pYZ2N respectively. Those strains were induced to express proteins as described in Materials and Methods. GFP, which can reflect the APOBEC3G's level in cells, was observed under fluorescent microscopy In addition, cells were harvested, lysed and analyzed with Western Blot (Fig. 3 B). When Vif expression was enhanced, the fluorescence in cells which can reflect the proteins' level of GFP-APOBEC3G was decreased (Fig. 3 A). In the top panel of Fig. 3 A, the intensity of green fluorescent dots were strongest which reflect the original level of APOBEC3G protein when expressed without Vif protein; in the middle panel, the intensity of green fluorescent dots were lower comparing to that in the top panel, which reflect that GFP-APOBEC3G protein's level was decreased with the expression of Vif protein; in the lowest panel, the intensity of green fluorescent dots were weakest and dispersed in the cells, which reflects that GFP-APOBEC3G protein's level was decreased most significantly when Vif protein was over-expressed. Similarly, Western Blot results showed that GFP-APOBEC3G protein level was significantly decreased when Vif protein was over-expressed (Fig. 3 B, lane 6) comparing to when Vif protein was normally expressed (Fig. 3 B, lane4) or not expressed (Fig. 3 B, lane 2).
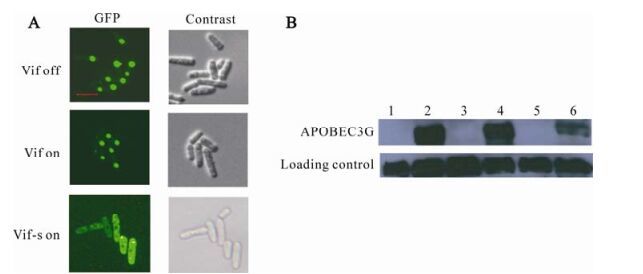
Figure 3. Vif mediates the degradation of APOBEC3G in S. pombe cells. pYZ3N-APOBEC3G and pYZ2N-vif-s were co-transformed into one S. pombe cells using lithium acetate method. A: Single colony were selected and protein expression were induced for 24 hours by diminishing the thiamine in the media (-T). Living cells were observed under fluorescent microscopy. In the experiments, strains containing pYZ3N-APOBEC3G and pYZ2N, strains containing pYZ3N-APOBEC3G and pYZ2N-vif were used as controls. The scale bar represents 10µm. B: Strains containing pYZ3N-APOBEC3G and pYZ2N, strains containing pYZ3N-APOBEC3G and pYZ2N-vif, strains containing pYZ3N-APOBEC3G and pYZ2N-vif-s were cultured in EMM media with(+T) or without(-T) thiamine. Expressed proteins were induced for 48 hours before the cells were harvested and lysed. The same quantity of proteins were subjected to electrophoresis and immunoblotting analysis, proteins were tested with anti-GFP and anti-tubulin antibodies. Lane 1: pYZ2N pYZ3N-APOBEC3G/SP223 +T; Lane 2: pYZ2N pYZ3N-APOBEC3G/SP223 –T; Lane 3: pYZ2N-vif pYZ3N APOBEC-3G/SP223 +T; Lane 4: pYZ2N-vif pYZ3N-APOBEC3G/SP223 –T; Lane 5: pYZ2N-vif-s pYZ3N-APOBEC3G /SP223 +T; Lane 6: pYZ2N-vif-s pYZ3N-APOBEC3G /SP223 –T.
Subcellular localization of HIV-1 Vif and APOBEC3G
Codon optimalization of vif gene enhances the expression of Vif protein
Vif mediates the degradation of APOBEC3G in fission yeast
-
Results of this study demonstrate that (ⅰ) the Vif protein of HIV-1 is localized mainly in the nuclei in fission yeast cells, (ⅱ) the APOBEC3G protein is concentrated at punctate bodies which are localized in cytoplasm and (ⅲ) Vif protein expression levels can be significantly enhanced through codon optimization. When over-expressed, a small portion of GFP-Vif showed cytoplasm localization in fission yeast. When co-expressed in same cells, over-expressed Vif protein can mediate the degradation of APOBEC3G in fission yeast cells, which suggests that both proteins may have similar functions in both mammalian cells and fission yeast cells. Taken together, these results indicated that fission yeast cells can be used to study the relationship between Vif and APOBEC3G effectively, but high levels of Vif protein expression are crucial for studying the effects of Vif function on APOBEC3G.
Our live imagingstudies revealed that GFP-Vif localized predominantly to the nucleus of fission yeast cells. Regarding the subcellular localization of the Vif protein, previously reported data has been contradi-ctory. Some reports showed that Vif localized predo-minantly in the cytoplasm and wasonly observed in modest amounts in the nucleus, a phenotypethat was sometimes attributed to overexpression (11, 15, 31). However, similar to feline immunodeficiency virus Vif (3), some data showed that Vif localized in the nucleus when observed in living cells, and the cytoplasm localization was ascribed to the methods of fixation and staining (26). In this study, the results demonstrated that a more substantial fractionof Vif localizes to the nucleus, and only a small part is localized in the cytoplasm when over-expressed through codon optimization, which is consistent with previous findings that show Vif interactswith nuclear proteins Sp140(19) and ZIN (7).
In this study, we explored a new S. pombe system for studying subcellular localization and functions of APOBEC3G, which will be a valuable tool for exploring the mechanism of APOBEC3G functions in cells. APOBEC3G has been shown to localize throug-hout the cytoplasm and to concentrate within punctate cytoplasmic bodies (36). In this report, we also found that APOBEC3G was mainly concentrated within punctuate cytoplasmic bodies and seldom distributed in the cytoplasm, suggesting that S. pombe is suitable for studying functions of APOBEC3G in cytoplasmic bodies. Our emphasis was on APOBEC3G because of its role in inhibiting Vif-deficient HIV-1 virus; its normal function and functional mechanism was not extensively studied. However, understanding the mechanism by which APOBEC3G functions in normal cells is important exploring the mechanisms by which APOBEC3G inhibits Vif-deficient HIV-1 virus and for the development of new anti-HIV-1 strategies based on APOBEC3G.
Codon optimization is a useful strategy for high level expression of proteins, especially for those proteins that are difficult to express. Vif is notoriously difficult to express autonomously in the absence of the viral Tat and Rev proteins. It has been shown that Rev facilitates HIV RNA export by binding to the RRE and by simultaneously interacting with the CRM1/Ran complex, which in turn interacts with components of the nuclear pore complex to mediate the energy-dependent translocation of the RNA molecule into the cytoplasm (16). In addition to RRE, numerous studies have indicated that some other factors were responsible for nuclear retention of viral mRNA in the absence of Rev (2). Regions of high AU content (13, 20, 30) as well as AUUUA motifs (29), collectively referred to as cis-acting inhibitory elements(INS), have been identified and largely account for the nuclear retention of unspliced and singly spliced HIV-1 mRNAs. However, in our study, although we didn't decrease the content of AU and AUUUA level in the gene, we still demonstrated that codon optimization of the vif ORFs led to a dramatic enhancement of protein synthesis in the absence of the viral regulatory proteins Tat and Rev. So in S. pombe cells, the structure of mRNA and the codon contains in the genes may be more important than the AU and AUUUA levels.
In this study, it was demonstrated that the degradation of APOBEC3G was enhanced by over-expression of Vif protein in a fission yeast system. It has been shown previously that the HIV-1 Vif protein was able to inhibit the APOBEC3G protein in a E. coli system (28). As far as we know, this is the first time anyone has reported that the Vif protein was able to induce the degradation of APOBEC3G protein in a S. pombe system. S. pombe is a relatively simple mono-cellular eukaryote which has been proved to be useful for studying as a model system with sophisticated laboratory operation techniques. For this reason S. pombe has been frequently used for the study of basic biological pathways and protein functions. In previous studies, S. pombe has been used as a model system to study functions of proteins of human cells or viral cells, including the study of the HIV-1 Vpr protein (40), and shows nearly the same phenotype as human cells. So demonstrating the use of S. pombe for the study of the Vif-APOBEC3G interaction suggest further applications for the study of Vif function and screening of suppressors of Vif function.
In summary, our studies suggest that the subcellular localization of HIV-1 Vif and human APOBEC3TG proteins in S. pombe mimic those found in human cells. In addition, our results further suggested that Vif mediated APOBEC3G's degradation in S. pombe cells is similar to degradation in human cells. Since S. pombe is a relatively simple monocellular eukaryote, S. pombe provides a useful model system to study genetic and biochemical aspects of HIV-1 vif gene functions.

 DownLoad:
DownLoad: