











-
Fibroblast growth factor (FGF) was originally identified in the mid of 1970s in the pituitary of brain extracts and possesses mitogenic activity for murine fibroblasts. The genes of FGF family have been identified in all metazoan from nematode to mice and humans but not in bacteria or yeast, highlighting its importance in a variety of multicellular biological processes such as mitogenesis, angiogenesis, cell proliferation, differentiation and cell migration (12). The role of FGF in many of these processes is conserved in a variety of organisms, while inappro-priate expression of some fgfs may lead to cancer (13, 24). FGFs can bind heparin or heparin sulfate proteoglycans to form oligomers. This complex interacts specifically with FGF receptors (FGFRs), a group of transmembrane tyrosine kinases that are activated upon FGF binding leading to receptor dimerization and autophosphorylation. The activated FGFRs then stimulate the signal transduction pathways (24). It is notable that an fgf homologue (vfgfs) has been identified in baculoviruses, but not in other viruses and microorganisms so far (23).
The vfgf is present in all baculoviruses that infect lepidoptera, whose genomes have been sequenced so far (1, 20), suggesting that vfgf plays an essential role in the infection cycle of lepidopteran baculoviruses. It has been demonstrated that both Autographa califor-nica nucleopolyhedrovirus (AcMNPV) and Bombyx mori NPV (BmNPV) encode functional vFGF, which share some conservative properties common to better characterized vertebrate and nematode FGF, such as structural features, heparin affinity and chemotatic function (6). Moreover, BmNPV vfgf is involved in efficient virus production in BmN cells and B. mori larvae (17); however, the AcMNPV lacking vfgf recombinant virus has no obvious effect on virus production and protein or DNA synthesis relative to wild type virus both in vitro and in vivo (7). These results imply that the vfgf has functionally differen-tiated during viral evolution even in those baculovi-ruses with close relative, such as AcMNPV and BmNPV belonged to group Ⅰ NPV. So, it will be very interesting to analyze the vfgfs from baculovirus with evolutional distance, such as Helicoverpa armigera NPV (HearNPV) of group Ⅱ NPVs.
HearNPV contains a DNA genome of 135 kb encoding 135 open reading frames (ORFs) of 50 amino acids (aa) or more and ORF113 encodes a putative vfgf homolog (3). In this study, we carried out a detailed transcriptional analysis, evaluated some biochemical characteristics of vFGF compatible with the function of other known FGFs, and assess its properties in the specific chemoattraction of cells.
HTML
-
HearNPV strain G4, HaCXW1 and HearNPV bacmid HaBacHZ8 was propagated in Hz-AM1 cell and H. armigera larvae (3, 5, 28). Hz-AM1, Sf9 and Se-UCR cells were cultured in Grace medium (Invitrogen) supplemented with 10% fetal bovine serum (Gibco-BRL) at 27℃; 293 and HepG2 cells were cultured in 1640 medium (Invitrogen) supple-mented with 10% fetal bovine serum (Gibco-BRL) at 37℃. H armigera hemocytes were isolated from hemolymph and resuspended in ice-cold anti-coagulant buffer (28) and immediately used for transmigration assays.
HearNPV BVs were purified from hemolymph of H. armigera fourth-instar larvae infected with HearNPV polyhedra according to IJkel et al (11). ODVs were purified from HearNPV G4 polyhedra derived from infected H. armigera fourth-instar larvae as described previously (3). The purity and integrity of BVs and ODVs were checked by electron microscopy.
-
The complete vfgf was amplified by PCR from HearNPV genome DNA by using the primer F1 (5'-GGGGAATTCATGTTTTCGGTATTGTTGAGA-3') and F2 (5'-GGGAAGCTTTTAGACATTATTTTTT TTTGA-3') and cloned into pGEX-KG and pMal-c2x (NEB, UK). vFGF-GST and vFGF-MBP fusion protein were expressed in Escherichia coli DH5α and TB-1, respectively. Purified vFGF-GST was used as an antigen to produce antiserum in rabbits (αHaFGF), the specificity of the antibodies was assessed by immuno-blotting (data not shown). The soluble vFGF-MBP fusion proteins were purified by Amylose resin column (NEB, UK) and used for chemotactic activity assay.
-
1×106 Hz-AM1 cells were infected with Hear NPV-G4 BV at an MOI of 10 and harvested at 0, 18, 24, 48, 72 and 96 hour post-infection (hpi). Total RNA was isolated from cells with Trizol (Gibco-BRL) according to the manufacture's guidelines. RNA was dissolved in 50 μL water and quantified by optical density measurements. Hz-AM1 RT-PCR was performed using 1 μg total RNA as template for each time-point. First-strand cDNA synthesis was performed with AMV reverse transcriptase (Promega) and a 15 nt-oligo (dT) primer. cDNA mixtures were amplified by PCR using primers F1 and F2 (9).
Cells were harvested at different time point after infection and lysed in SDS-PAGE loading buffer. Uninfected cells were taken as a control. Proteins were separated by SDS-PAGE and blotted onto Hybond-N membranes (Amersham). Duplicate blots were prepared and processed for immunoblotting. HearNPV vFGF-specific antiserum (αHaFGF) and alkaline phosphatase-conjugated goat anti-rabbit im-munoglobulin (Sino-American) were used as the primary (dilution with 1:1 000) and secondary antibo-dies (dilution with 1:1 000), respectively. The signal was detected using a BCIP/NBT kit (Sino-American) (9).
-
Electron microscopy (EM) was used to examine the integrity of virions with 2% of phosphotungstic acid (PTA) staining. For immunogold labeling, purified virions were loaded onto a collodion-coated EM grid for 12 h. After the removal of excess sample solution, a HearNPV vFGF-specific antibody (αHaFGF, dilution with 1:500) was added onto the grid and incubated for 1.5 h at room temperature. Following three wash steps in PBS for 5 min per wash at room temperature, grids were incubated with 15 nm gold conjugated anti-rabbit IgG (dilution with 1:25) for 1.5 h. After three 5-min washing steps in PBS, the samples were stained with 2% PTA for 2 min, drained, and examined under the EM.
-
1×106 Hz-AM1 cells were infected with HearNPV-G4 BV at a MOI of 10 pfu/cell. At 72 hpi the infected culture supernatants were harvested and centrifuged at 24 000 r/min for 90 min to remove budded viruses. The resulting supernatants were concentrated with Amicon Ultra-15 filter devices (10 000 NMWL me-mbrane, Millipore) at 5 000×g for 30 min and diluted in 15 mL binding buffer (10mmol/L sodium phosphate, pH7). The infected Hz-AM1 cells were pelleted, resuspended in binding buffer and then lysed by sonication. Subsequently, these samples were loaded respectively onto ac1-mL HiTrap affinity column (Amersham) and washed with phosphate buffer (10 mmol/L NaH2PO4) containing different concentrations of NaCl from 0.2 to 2.0 mol/L according to the manufacture's manual. Each fraction was collected and proteins were detected by immunoblotting.
-
Cell migration was assessed in Costar transwell trays with polycarbonate membrane inserts (6, 7) Five times 104 Hz-AM1, Sf9, SeUCR, Ha hemocyte, 293 and HepG2 cells were pelleted, washed three times with PBS and resuspended in PBS, and then loaded onto 8 μm pore-size transwell inserts, which were then transferred to the plates containing different amounts of purified proteins or virions. After 1.5 to 2 h incubation at 27℃ (or 37℃ for mammalian cells), transwell inserts were removed and the number of cells migrated to the membrane was measured by microscopic examination. The chemotactic index is define as the ratio of the number of cells between the treatment with protein/virion and the control with MBP/TE (The cut-off is 2.0), the chemotactic index exceeds the cut-off 2.0 means that protein/virion has chemotactic activity. All experiments were performed independently at least three times.
For antibody neutralization analysis, 3.5×108 pfu/ mL HaCXW1 (4) were pre-incubated with antiserum αHaFGF in a ratio (BV/antiserum, V/V) of 1:1, and 1:2 for 2 h at 37℃. The αHa98, the antiserum to HearNPV ORF98 or per os infectivity factor 3 (PIF-3) (stored in lab) was used the negative control. The titres of the three antibodies are 1:5 000 with Western blot assay. Then, neutralized HaCXW1 was used for cell migration.
Virus and cells
Expression of vFGF and generation of anti-vFGF antiserum (αHaFGF)
Transcription and translation analysis
Immunogold electron microscopy (IEM)
vFGF affinity to heparin
Chemotactic activity assay
-
The HearNPV ORF113 encodes a FGF homolog, with a predicated size of 33.5 kDa (3). Sequence analysis indicated that the vFGF contains a 23 aa-long typical signal sequence in the N terminus and the conserved domains of the FGF family (2, 26), in-cluding FGF heparin binding sites and FGF receptor interaction sites. Computational analysis showed that vFGF could form 12 strands in the conserved region. As compared to AcMNPV and BmNPV vFGF, the C terminus of HearNPV vFGF has an additional 114 amino acids (aa). Four putative N-glycosylation sites, one N-myristoylation site, three protein kinase C phosphorylation sites and one Casein kinase Ⅱ phos-phorylation site were predicated by sequence analysis.
-
Transcription of HearNPV vfgf was examined by RT-PCR, using total RNA isolated from Hz-AM1 cells infected with HearNPV G4. A single 900 bp band was first detected at 24 hpi and continued to be present until 96 hpi indicating that the vfgf is a late gene and probably uses the late transcription initiation codon of TAAG, 18 nt upstream of the translation start codon ATG. (Fig. 1A). The predicted size is in agreement with the size of the detected transcript.
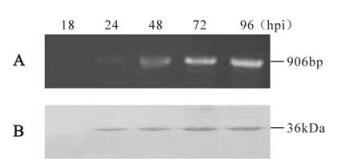
Figure 1. Transcription and expression analysis of the vfgf. A: Total RNA was extracted at different time-points (hpi) from Hz-AM1 cells infected with HearNPV at an MOI of 10 pfu/cell. Reverse transcription was performed using an oligo (dT) primer and PCR amplification was done using the specific primers. M, DNA marker DL2000. B: Western blot analysis of the vFGF in HearNPV infected Hz-AM1 cells from 18 to 96 hpi. The vFGF was identified using polyclonal antiserum (αHaFGF) and detected with alkaline phosphatase-labeled secondary antibody goat anti-rabbit immunoglobulin (Sino-American).
During infection, HearNPV vFGF antiserum specifically detected a 36 kDa polypeptide (Fig. 1B). This band first appeared at 24 hpi and continued be present at 96 hpi. The size of 36 kDa is a little larger than the predicted 33.5 kDa, which may be the consequence of post-translational modifications such as glycosylation, which has also been shown for BmNPV vFGF (15). From the results of transcription and translation analysis, we concluded that the vfgf is most likely a late gene, which is different from what has been observed for BmNPV and AcMNPV (6, 18).
-
Hydropathy plot analysis showed the existence of the typical signal sequence in the N terminus of vFGF (1-23 amino acids) suggesting that the vFGF might be a secreted protein. Western-blot analysis indicated that a 36 kDa protein could be detected in the infected culture supernatants (data not shown). In addition, it is notable that the 36 kDa protein could also be detected in HearNPV BVs, but is absent in ODVs (Fig. 2A), indicating that the vFGF is associated with HearNPV BVs. Immune colloidal gold results indicate that the vFGF is associated with BVs envelope and not on ODVs (Fig. 2B C).
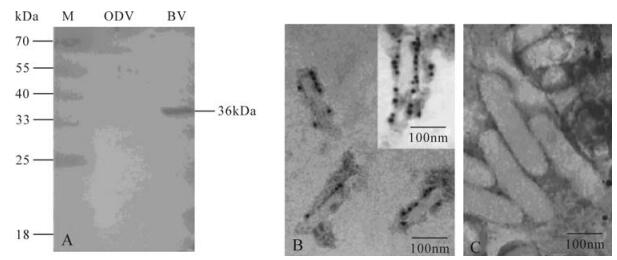
Figure 2. Western blot analysis of vFGF in purified BVs and ODVs (A). About 10 μg of purified BVs and ODVs were separated by SDS-PAGE and analyzed by Western blot using polyclonal antiserum (αHaFGF) as the first antibody and detected with alkaline phosphatase-labeled secondary antibody goat anti-rabbit immunoglobulin (Sino-American). Electron micrograph of immunogold labeled purified HearNPV BVs (B) and ODVs (C). BVs and ODVs were probed using αHaFGF counterstained with gold spheres coupled to anti-rabbit IgG.
-
One property of FGFs is its ability to induce cell motility. The recombinant vFGF protein was expressed and purified in E. coli. As expected, the recombinant vFGF has clear chemotactic activity on Hz-AM1 cells, but neither other tested insect cells such as Sf9 and Se-UCR, nor mammalian cell lines such as 293 and HepG2 (Fig. 3A). The chemotactic activity on Hz-AM1 increased with the protein concentration in-creasing (from 0.05 to 1 μg/mL). When the protein concentration reaches 1 μg/mL, the chemotactic activity is maximal and the chemotactic index is 4.46. However the chemotactic activity declined when the protein concentration surpassed 1.0 μg/mL, probably due to diffusion of vFGF and preventing a gradient of FGFs from being established. This result showed that the HearNPV vFGF has specific chemotactic activity on the Hz-AM1 cell line.
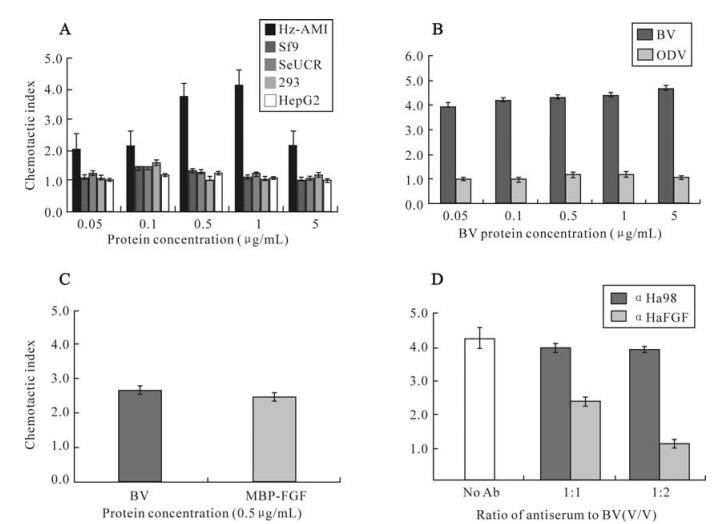
Figure 3. vFGF chemotactic activity assay. A: Chemotactic activity of the recombinant FGF on Hz-AM1, sf9, SeUCR, 293 and HepG2. Results are given as the mean±s.e.m. of the chemotactic index, which is the ratio of the number of cells migrated to the membrane between MBP-FGF and MBP treatments. B: Chemotactic activity of purified ODVs and BVs on Hz-AM1. Data are expressed as chemotactic index, which is the ratio of the number of cells migrated to the membrane between virions and 0.1×TE treatments. C: MBP-FGF and HearNPV BVs chemotactic activity on H armigera larvae hemocyte. Data are expressed as chemotactic index, which is the ratio of the number of cells migrated to the membrane between MBP-FGF/BVs and MBP/0.1×TE treatments. D: Chemotactic activity of BVs neutralized by antiserum αHaFGF. The bars above each column indicated the mean ± standard error of at least three independent experiments.
Since vFGF is associated with BVs, we investigated BV chemotactic activity of BVs on Hz-AM1 cells. Fig 3B shows that BVs have clear chemotactic activity on Hz-AM1 cells when the concentration of BV ranged from 0.05 to 5.0 μg/mL, whereas the ODV chemotactic index fluctuated between 0.992 and 1.263. So BV has clear chemotactic activity on Hz-AM1 cells but ODV shall not. This result is in agreement with the association of vFGF with BV and not with ODV. Furthermore, both HearNPV BVs and recombinant vFGF (both at 0.5 μg/mL) were able to stimulate migration of hemocytes obtained from last instar H. armigera larvae, implying that the vFGF is able to attract hemocytes in the infected host (Fig. 3C). In contrast, AcMNPV BVs did not show any chemo-tactic activity on Sf9, HzAM1 or SeUCR (data not shown).
To further determine whether the chemotactic activity of HearNPV BV was due to vFGF, BVs (0.5 μg/mL) were preincubated with αHaFGF before the chemotactic activity assay. The chemotactic activity declined by approximately 50 percent when HearNPV BVs were neutralized by αHaFGF at a ratio (an-tiserum/BV, V/V) of 1:1. When the antiserum ratio increased to 2:1, BVs completely lost chemotactic activity on Hz-AM1 cells (Fig. 3D). These results clearly indicated that the vFGF, whether or not associated with BV envelope, is the chemotactant.
-
FGFs bind heparan sulfate proteoglycans in the extracellular matrix, and upon cleavage, complexes of heparin-bound FGFs bind and activate FGFRs. Previously characterized FGFs bind heparin-Sepharose and elute at high salt concentrations (13). To test whether vFGF had high affinity to heparin, the supernatant without BVs and the cellular lysate were collected from HearNPV infected Hz-AM1 cells applied to heparin-Sepharose; the bound proteins were eluted with increasing concentrations of NaCl. FGF effective elution concentration is between 1.6 and 1.8 M NaCl (Fig. 4). The result indicated that HearNPV vFGF also has the property to strongly bind heparin, which is critical for the function of all FGFs (13, 24). In addition HearNPV vFGF is a secreted protein and may be glycosylated. On the other hand, the recombinant FGF fusion protein could also bind heparin (Fig. 4) indicating that the glycosylation of vFGF is not essential for its chemotatic activity.
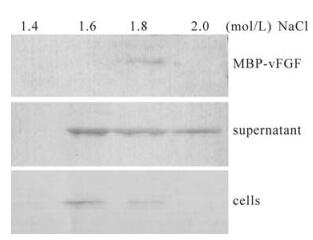
Figure 4. vFGF affinity to heparin. The supernatant and the cells were collected from HearNPV infected Hz-AM1 and MBP-FGF fusion proteins applied to heparin-Sepharose, respectively, and bound proteins were eluted with increasing concentrations of NaCl (0.2 to 2.0 mol/L). The eluent was loaded per lane, separated in a 12% gel, transferred onto a Hybond membrane and analyzed by Western blot analysis using polyclonal antiserum (αHaFGF) and detected with alkaline phosphatase-labeled secondary antibody goat anti-rabbit immunoglobulin (Sino-American).
Alignment of FGF sequences and structure analysis of vFGF
Transcriptional and translational analysis
Immunodetection of the vFGF protein in BVs and ODVs
Chemotaction properties of vFGF
vFGF affinity to heparin
-
Fgf genes have been identified in a variety of organisms and have very important functions in their biological processes (23). However, in viruses, fgf homologues have only been identified in bacu-loviruses and they must have a critical role in infection. In this paper, we characterized the HearNPV vfgf from group Ⅱ NPV. Transcriptional and translational analysis indicated that the vfgf most likely is a late gene and probably uses the late transcription initiation TAAG in the promoter region. 5' RACE should indicate whether this is the case or not. Western blot analysis detected a 36 kDa protein during infection and in BVs, but not in ODVs. The actual size is slightly larger than the predicted size (33.5 kDa) based on the nucleotide sequence. This suggests that the vFGF might undergo post-translation modification upon synthesis.
It has been reported that vFGFs in both Spodoptera litura NPV and BmNPV are glycosylated (15), while the AcMNPV vFGF with a signal peptide is not glycosylated and is not secreted (7). We found that the glycosylation sites at positions 44 (Asn) of BmNPV vfgf, which is crucial for vFGF secretion (15), was also conserved in HearNPV vFGF (N49), but not in AcMNPV vFGF, strongly implying that the HearNPV vFGF may undergo N-glycosylation. Western-blot analysis (Fig. 2) confirmed the vFGF is secreted into the supernatant of HearNPV-infected Hz-AM1 cells, which is similar to BmNPV vFGF (15).
Interestingly, vFGF can also be detected in HearNPV BVs but is absent in ODVs, indicating that the vFGF is not only a secreted protein but also is associated with HearNPV BVs. The immune colloidal gold analysis further confirmed that the vFGF is localized on the envelope of BVs and not on ODV (Fig. 2B C). BV also possesses chemotactic activity on Hz-AM1 cells (Fig. 3B), but this is not for ODVs. Whether vFGF is really structural component of BVs or is adhered to it remains to be determined.
The HearNPV vFGF shows special chemotactic activity on Hz-AM1 cells but not on other tested cell lines, while the AcMNPV vFGF was able to stimulate transmigration of Sf-21 and TN-368 cells (6). Those results suggest that the HearNPV vFGF chemotactic activity is cell specific. The reason may be that HearNPV and AcMNPV vFGF recognize different FGF receptors. Recently, Susumu et al have identified BmBtl and SfBtl as the receptors for BmNPV vFGF (16). Fibroblast growth factors comprise a structurally relatedfamily of 22 molecules (21, 30). While FGF 1 appears to be a universal FGFR ligand and activate all FGF receptors, no other FGF ligand could activate all FGFRs (22, 30). The cellular specificity of vFGFs' activity might also contribute to the viral host range. It has been reported that AcMNPV with a wide host range can replicate in different lepidopteran cell lines, but can not replicated in a cell line from H. zea (14). On the other hand, HearNPV with a narrower host range can only infect the cell lines from H. spp. However, the relation between and viral host range need to be further clarified.
It has been reported that the hemocytic and tracheal systems play a central role in baculovirus patho-genesis by amplifying virus and disseminating in-fection during the baculovirus secondary infection (8, 27, 29). vFGF may be involved in disseminating infection systemically. After virus initially infect the midgut, the secretive vFGF and BV with vFGF could chemotact hemocytes to the infected focus to accelerate virus dissemination and to establish secondary infection. The latest studies indicate that Bmbtl, the receptor of both the BmNPV vFGF and the cellular FGF, was abundantly expressed in the trachea and midgut in B. mori larvae, while moderate expression observed in the hemocytes and the B. mori cultured cell line BmN (16). The Drosophila FGF gene, branchless, mainly controls the migration and differentiation of tracheal cells (25). And the brea-thless receptor is mainly expressed at tracheal surface during tracheal system branch morphogenesis (10, 19).

 DownLoad:
DownLoad: