









HTML
-
Bacteriophages (phages) are viruses that specifically infect and kill bacteria. They are ubiquitous throughout all environments that bacteria inhabit. Following their discovery by F.W. Twort in 1915 and F. d'Herele in 1917, bacteriophages were recognized as potential agents to treat bacterial diseases and phage therapy has been used in clinical practice ever since. However, with the invention of penicillin and other antibiotics, phage therapy was largely given up by most Western countries, with the exception of Russia, Georgia and Poland. Recently, the increasing incidence of antibiotic resistance and untreatable bacterial infections has led researchers to repurpose these bacteria-specific viruses as important agents against pathogenic bacteria, in human and veterinary medicine, food industry, and agriculture.
Klebsiella pneumoniae is a facultative anaerobic, gram-negative, rod-shaped bacterium that is very common in hospital environments. This organism is an important causative agent of nosocomial and opportunistic infection in humans, such as pneumonia, urinary tract infections, bacteremia, and intra-abdominal infections (Podschun and Ullmann, 1998; Fang et al., 2007). Infections caused by K. pneumoniae are frequently life threatening in infants, the elderly and immunocompromised individuals (Ko et al., 2002). In addition, extensive use of broad-spectrum antibiotics in hospitalized patients has facilitated the development of multidrug-resistance due to the production of extended-spectrum β-lactamases (Lin et al., 2007). The declining efficacy of antibiotic therapies leads to the necesssary development of novel therapeutics for K. pneumoniae infections (Kumari et al., 2011; Hung et al., 2011). The goals of this study were to isolate phages that were lytic against K pneumoniae isolates, and characterize their biological features to evaluate their potential as therapeutic agents to fight K. pneumoniae infections.
Here 25 clinical isolates of K. pneumoniae, previously isolated at Department of Clinical Laboratory, the third affiliated hospital of Jilin University, were used for phage isolation and identification (Table 1). Phage isolation was conducted as described previously (Yang et al., 2010; Parasion et al., 2012). Purified phage particle morphology was examined by transmission electron microscope (TEM) of negatively stained preparations. The extracted phage DNA was subjected to restriction digestion with nine restriction enzymes (EcoR Ⅰ, EcoR Ⅴ, Xho Ⅰ, Nde Ⅰ, Pst Ⅰ, Bgl Ⅱ, Hind Ⅲ, Xba Ⅰ, and Not Ⅰ) and analyzed by agarose gel electrophoresis. Multiplicity of infection (MOI) for K. pneumoniae was determined by mixing bacteria and phages at ratios ranging from 10-4 to 102 PFU/CFU in tubes containing 108 CFU/mL K. pneumoniae. The tubes were incubated at 37 ℃ for 4 h, and the number of plaques on double-layer solid medium was calculated. One-step growth experiments were carried out according to a standard method (Hyman and Abedon, 2009).
Bacterial species, strain tested Sensitivity to φKp-lyy15* Klebsiella pneumonia K1, K2, K15, K17, K18, K19 + K31, K36, K62, K67, K81 + K3, K5, K6, K9, K11, K12, K13 – K20, K21, K22, K26, K27, K28, K30 – Escherichia coli ATCC 25922 – Pseudomonas aeruginosa ATCC 27853 – *: Strains were grown in LB medium at 37 ℃. Spots were divided into two categories: Clear zone (+) and no reaction (–). Table 1. Lytic activity of φKp-lyy15 against various Klebsiella strains.
To determine the proteins of phage particles, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed as previously described (Boulanger, 2009). Resistance to different pH values at 37 ℃ was carried out as previously documented (Merabishvili et al., 2014). Phage thermal stability of phage at different temperatures (50, 60, 70 and 80 ℃) was determined by incubating the phage (107 PFU/mL) at the indicated temperature for 30 and 60 min in nutrient broth (pH 7.0); and then calculating the surviving phages. The host rang of isolated phages was measured using spot test on 25 K. pneumonia clinical strains, Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853.
This study describes a newly isolated bacteriophage φKplyy15 specifically infecting K. pneumoniae isolates and its biological features. The Klebsiella bacteriophages described in publications belong to Podoviridae, Myoviridae and Siphoviridae families (Ackermann, 2001; Kumari et al., 2010). Our results showed that phage φKp-lyy15 has an icosahedral head 50 nm in diameter and a filamentous tail 120 nm in length. Thus, the bacterial virus was classified as a representative of Siphoviridae families (Figure 1A and 1B). The phage genome is presented by double-stranded DNA that is digested with the restriction endonucleases EcoR V, Nde Ⅰ, and Bgl Ⅱ (Figure 1C), and our future research will focus on further detailed analyses of the entire φKp-lyy15 genome to understand the genetic characteristics of this bacteriophage. As shown in Figure 1D, the group with an MOI of 10-3 yielded the highest production of phage progeny (5 × 109 PFU/mL). One-step growth tests were carried out to determine the latent time and phage burst size of φKp-lyy15. The latent period, defined as the time interval between the adsorption and the beginning of the first burst, was 10 min, and the burst size was 65 PFU/infected cell (Figure 1E). In this study, the burst size was equivalent to the number of phage at plateau phase (6.5 × 109 PFU/mL) divided by the initial number of host cells (1.0 × 108 CFU/mL).
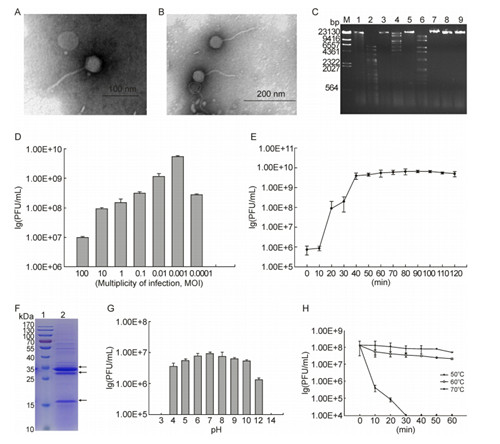
Figure 1. (A, B) Morphology of phage φKp-lyy15 as revealed by TEM. (A)Scale bar=100nm, (B)Scale bar=200nm. (C) Restriction of phage genomic DNA. M: DNA marker; Lanes 1–9: Digestion of DNA with EcoR Ⅰ, EcoR Ⅴ, Xho Ⅰ, Nde Ⅰ, Pst Ⅰ, Bgl Ⅱ, Hind Ⅲ, Xba Ⅰ, and Not Ⅰ respectively. (D) The MOI experiment. (E) One-step growth experiments. Latent phase from 0 to 10 min; Rise phase from 10 to 40 min, and Plateau phase from 40 to 120 min. (F) SDSPAGE of phage protein. Lane 1: PageRuler Prestained Protein Ladder #26617 (Thermo), Lane 2: Phage φKplyy15 (109 PFU/mL), arrows indicate major protein bands. (G) pH stability tests of phage φKp-lyy15. (H) Thermal stability tests of phage φKp-lyy15.
Purified phage particles were subjected to SDS-PAGE to quantity the number of structural phage proteins. Three major protein bands and at least 12 minor protein bands were detected, with molecular weights ranging from approximately 15 to 170 kDa (Figure 1F). The most predominant polypeptide band of approximately 35 kDa presumably corresponded to a major capsid protein. φKp-lyy15 stability was investigated at different pHs and temperatures. The results showed that no apparent effect on φKp-lyy15 was observed after 1 hour of incubation at pH values ranging from 5 to 10. However, the phage dramatically lost its activity at pHs lower than 3 and higher than 13 (Figure 1G). The thermal stability test shown in Figure 1H indicates that phage φKp-lyy15 was relatively heat stable, the 5.1 × 107 PFU/mL and 2.1 × 107 PFU/mL titers of active phages were detected after 60 min incubation at 50 ℃ and 60 ℃, respectively. Furthermore, the phage titer quickly dropped at 70 ℃, and no active phages were detected after 30 min.
The main aim of this study was the identification of a lytic bacteriophage with potential for therapeutic use. Therefore, the phage's lytic activity and host specificity were tested against 25 strains K. pneumonia strains. The phage was shown to specifically infect and lyse 44% (11 of 25) of K. pneumonia strains by forming clear zones (Table 1). Two other gram-negative microorganisms, E coli and P. aeriginosa, were insensitive to the phage. In contrast, the phage KP36 belonging to Siphoviridae exhibited lower lytic activity against K. pneumonia strains (7%–15%) as documented in a recent publication (Kesik-Szeloch et al., 2013).
In conclusion, the lytic bacteriophage φKp-lyy belonging to the Siphoviridae family specific for K. pneumoniae was isolated and characterized. φKp-lyy displayed a short latent period, stability to a wide pH rang, high thermal resistance, and lytic activity toward a relatively broad range of K. pneumoniae isolates. Thus, phage φKplyy should be considered as a candidate for inclusion in phage cocktails to control K. pneumoniae-associated nosocomial infections.
-
This study was supported by the Jilin Industrial Technology Research and Development Projects (Grant number: 2013C015-2) and Jilin high tech industry development project. All of the authors declare that they have no competing interests. This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: