










HTML
-
The epidemic patterns of HIV-1 subtypes are continuously evolving, and subtypes B and C are dominant (accounting for approximately 60% of infections). Subtype B accounts for just a little more than 12% of infections worldwide and more than half of HIV infections worldwide are caused by subtype C (Geretti, 2006). Nevertheless, subtype B, found predominantly in the United States and Europe, is the most widely studied. Therefore, studies of subtype C should be a priority (Saxena et al., 2012). There are substantial differences between subtype B and C HIV, including sequence, structural, and antigenic variation (Saiyed et al., 2011), each of which can influence the biological properties of the virus, susceptibility to existing and candidate antiretroviral drugs, and evolution of antiretroviral drug resistance (Cohen, 2012). These differences also imply that the treatment approach for the subtype B virus may not be directly applicable to subtype C (Cohen, 2012; Saxena et al., 2012). In fact, subtype C HIV appears to be transmitted more efficiently than the other subtypes (Essex, 1999). Therefore, it is of great importance to examine differences in pathogenic potency among these subtypes to understand the genetic and functional characteristics of these viruses.
The HIV-1 trans-activator Tat protein is essential for virus replication and the progression of HIV infection. Therefore, understanding functional differences in Tat between subtype B and C will provide important insight into the differential pathogenesis. Substantial amino acid variation has been observed among the clade-specific Tat proteins. Differences in conserved amino acids between B-Tat and C-Tat may result in functional discrepancies. For example, C-Tat is less neurotoxic than B-Tat, probably as a result of alterations in the dicysteine motif within the neurotoxic region of B-Tat (Mishra et al., 2008). However, it is not clear whether there are other functional differences between B-Tat and C-Tat.
Tat is an 86-101 residue regulatory protein (9-11 kDa) produced early in HIV-1 infection; it regulates productive and processive transcription from the HIV-1 long terminal repeat (LTR) (Loret et al., 1992; Karn, 1999; Gatignol and Jeang, 2000). The viral Tat mRNA is composed of two exons. The first exon encodes 72 amino acids (residues 1-72, hereafter Tat72) and contains three important functional regions: the cysteine-rich region (amino acids 22-37), the basic region (amino acids 49-57), and the glutamine-rich region (amino acids 58-72). The second exon encodes a variable number of amino acids (residues 14-29) that contribute to viral infectivity and other functions (Rana and Jeang, 1999).
Tat72 is generated in the late stage of the HIV-1 infection cycle. The 86-residue truncated form of Tat (hereafter Tat86), produced early in HIV-1 infection, is generated due to a premature stop codon within the second exon. Though the full-length form of Tat (i.e., the 101-residue form, referred to as Tat101) is predominant in HIV-1 clinical isolates (Gibellini et al., 2005), Tat86 and Tat72 are used more widely for in vitro studies. Therefore, the extreme C terminus of Tat is not frequently examined, despite its demonstrated significance in several biological assays (Ott et al., 1997; Xiao et al., 1998; Smith et al., 2003; Campbell et al., 2005; Passiatore et al., 2009; Lopez-Huertas et al., 2010).
In this study, we compared the stability of Tat between subtype B and C and the ability of each to transactivate HIV-1 LTR and activate NF-κB. We also assessed the effect of truncated forms of Tat on stability, LTR expression, and NF-κB activity. We observed differences in stability between B-Tat and C-Tat; the latter was a significantly stronger transactivator of LTR expression and showed a greater ability to activate the transcription factor NF-κB. The carboxyl-terminal region of Tat is involved in regulating the stability and transcriptional activity of Tat.
-
NL4-3 Tat and 1084i Tat were amplified from the infectious molecular clone NL4-3 and an infectious molecular clone of a pediatric clade C strain, HIV1084i, which was isolated from a Zambian infant infected with HIV-1 (Grisson et al., 2004), by overlapping polymerase chain reaction (PCR). The PCR products were inserted into the BamHI-XhoⅠ sites of the mammalian expression vector pcDNA3.1 to create pcDNA3.1-Tat-Flag. The truncated forms, CTat86-Flag, CTat72-Flag, and BTat72-Flag were generated by insertion of the cDNA fragments into the pcDNA3.1 vector. The NL4-3 LTR was amplified from infectious molecular clone NL4-3, and then inserted into the KpnⅠ-XhoⅠ site of the pGL3-Basic Vector.
-
HEK293T cells were maintained in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS), penicillin (50 U/mL), and streptomycin (50 mg/mL) at 37°C in humidified air with 5% CO2. Transfection was performed using polyethyleneimine (1 mg/mL) (Sigma, St. Louis, USA) and corresponding plasmids. In brief, for each 24-well plate, 1.0 × 105 293T cells were seeded with DMEM containing 10% FBS for 20 h before transfection. Less than 1 μg of plasmid DNA was suspended in 100 μL of DMEM in a polystyrene tube, and polyethyleneimine (3 μL for 293T) was added and immediately mixed before incubation for 10 min at room temperature. Thereafter, the mixture was distributed gently onto the cell culture and incubated at 37 °C in humidified air with 5% CO2. The medium was exchanged with complete medium (DMEM supplemented with FBS and antibiotics) 8 h later.
-
HEK293T cells transfected with pcDNA3.1-Tat-Flag were treated 24-h post-transfection with medium containing 200 μg/mL cycloheximide (CHX) (Sigma-Aldrich). Cells were harvested at 0, 2, 4, 6, 8, and 10 h post-treatment, and western blotting was performed on lysates. Band (pixel) intensities were quantified using ImageJ and corrected for background levels by sampling an empty area. The mean natural log values of band intensities (from three independent experiments) were plotted against time. Tat-Flag half-life was determined mathematically by fitting values to the linear-transformed exponential-decay formula: ln(y) = ln(b) -at, where y is the Tat-Flag band intensity at time t, b is the initial Tat-Flag band intensity at 0 h, and a is the decay constant and is equivalent to the slope of the curve.
-
HEK293T cells grown in 24-well plate wells were co-transfected with 100 ng of pLTR-luc or 3κB-luc and 20 ng of pRL-SV40, with or without 200 ng of pcDNA3.1-Tat-Flag. The cell medium was replaced with fresh medium 6 h after transfection. After transfection for 48 h, luciferase activity was measured using the Steady-Glo Luciferase Assay System (Promega, Madison, USA). Luciferase activity was normalized to Renilla luciferase activity. The data are presented as means ± SD (n =3).
-
HEK293T cells transfected with CTat101-Flag, CTat86-Flag, CTat72-Flag, BTat86-Flag, and BTat72-Flag were treated with (+) or without (−) MG132. Total RNA was isolated from cells using TRIzol reagent (Invitrogen, Carlsbad, USA) according to the manufacturer's instructions. The isolated RNA was reverse-transcribed into cDNA using reverse transcriptase (Promega). Real-time PCR was performed using the SYBR Green PCR Kit (TAKARA, Kusatsu, Japan) and a Real-Time PCR Detection System (BIO-RAD IQ2; Hercules, CA, USA). A melting curve analysis was performed to ensure the amplification of a single product. The following primers were used: BTat-F, 5′-GACTAGAGCCCTGGAAGCATC-3′; BTat-R, 5′-CCATAGGAGATGCCTAAGGC-3′; CTat-F, 5′-CTAGAGCCCTGGAATCATCC-3′; CTat-R, 5′-CCATAGGAAATGCCTAAGC-3′; GAPDH-F, 5′-ATCACTGCCACCCAGAAGAC-3′; GAPDH-R, 5′-ATGAGGTCCACCACCCTGTT-3′.
Plasmid construction
Cell culture and transfection
Protein translation arrest
Luciferase reporter assays
Quantitative real-time PCR analysis
-
The half-life of Tat was studied by a CHX chase assay. CHX treatment has been used to determine the half-lives of several proteins, including Tat (Lata et al., 2015). HEK293T cells expressing Tat-Flag (Figure 1A) were treated with CHX and harvested at various time points. Western blotting was then performed on the cell lysates. As shown in Figure 1B, the half-life of NL4-3 Tat (1-86, B-Tat) was 2.8 h, in fair agreement with the results of Sivakumaran et al. (2009). However, the half-life of 1084i Tat (1-101, C-Tat) was 5.5 h, which was twice of NL4-3 Tat. In addition, we examined the stability of Tat in Jurkat cells. The half-life of 1084i Tat was longer than that of NL4-3 Tat, similar to the results for HEK293T cells (Figure 1C)

Figure 1. The different stabilities of NL4-3 Tat and 1084i Tat in HEK293T cells. (A) Schematic representation of Tat-Flag-expressing plasmids. The cDNAs of NL4-3 Tat-Flag and 1084i Tat-Flag were inserted into the BamHI-XhoⅠ sites of the mammalian expression vector pcDNA3.1 (+) to create pcDNA3.1-Tat-Flag-expressing plasmids. (B) HEK293T cells expressing Tat-Flag were treated with CHX and harvested at 0, 2, 4, 6, 8, and 10 h post-treatment. Western blotting was performed on the total-protein-equalized lysates. The actin levels show equivalent sample loadings. The Tat-Flag band intensities were quantified using ImageJ, and their natural log values were plotted as a function of time. The calculated Tat-Flag half-life is shown. The data points represent the means and standard deviations of three independent experiments. (C) The half-life of Tat-Flag was examined in Jurkat cells.
-
Given that 1084i Tat is 15 amino acids longer than NL4-3 Tat in the C-terminal region, we examined whether the C-terminal region of 1084i Tat affects stability. Figure 2A shows that the half-life of the truncated CTat86 was 3.8 h. The stability of CTat86 was slightly lower than that of C-Tat, but it was still more stable than NL4-3 Tat. We also observed that the half lives of BTat72 and CTat72 (the first exon form of Tat) were 1.6 h and 2.4 h, respectively (Figure 2A-2D). These results suggest that the carboxyl-terminal region truncation makes Tat fragile and sensitive to degradation. Collectively, these results demonstrate that the stability of B-Tat and C-Tat were both modulated by their carboxyl-terminal regions and C-Tat was more stable than B-Tat.
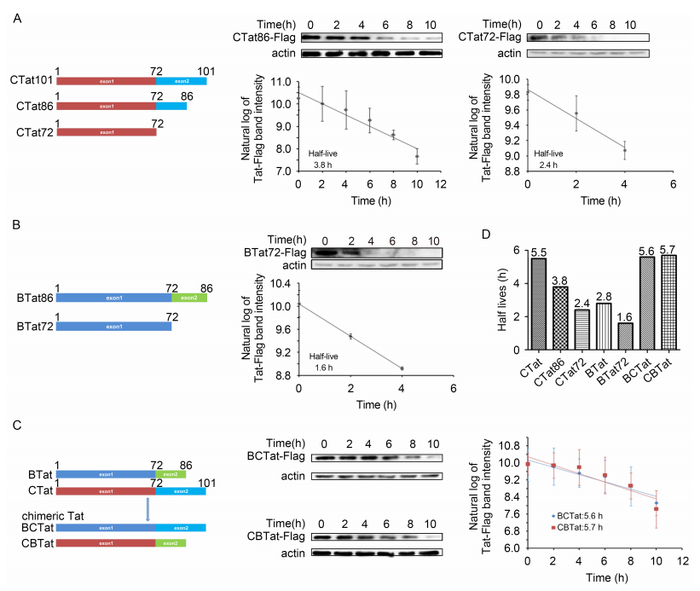
Figure 2. The carboxyl-terminal region of Tat regulates stability. (A, B, C) The half-lives of C-Tat, B-Tat, the carboxyl-terminal region deletion mutants, and chimeric Tat were determined. (D) Comparison of half-lives of different Tat mutants.
It is possible that the amino acid differences in the carboxyl-terminal region between NL4-3 Tat and 1084i Tat contribute to the observed differences in protein stability. We generated chimeric Tat proteins by swapping the clade-specific carboxyl-terminal regions (Figure 2C). Replacement of the NL4-3 Tat carboxyl-terminal region with the 1084i Tat carboxyl-terminal region resulted in enhanced stability of the chimeric BC-Tat (5.6 h) compared to B-Tat. However, the half-life of chimeric CB-Tat (5.7 h) was similar to that of C-Tat. This finding indicated that the specific domain of C-Tat is not responsible for the increased protein stability.
-
Tat is degraded by an ubiquitin-independent proteasomal pathway (Sivakumaran et al., 2009). HEK293T cells expressing Tat-Flag were treated with the proteasome inhibitor MG132, and this treatment substantially increased the stability of B-Tat and C-Tat (Figure 3A, 3D), clearly demonstrating that proteasome activity is required for Tat degradation. Moreover, for CTat101, MG132 treatment resulted in a significant increase (2.1-fold) in Tat-Flag compared to vehicle-treated controls, as assessed by western blotting (Figure 3B). The increases in CTat86, CTat72, BTat86, BTat72, BC-Tat, and CB-Tat by MG132 treatment were 3.8-fold, 4.5-fold, 4.1-fold, 6.4-fold, 1.8-fold, and 1.5-fold, respectively (Figure 3B and 3E). Based on a quantitative real-time PCR analysis, we found that the mRNA levels of Tat and its mutants did not change significantly after MG132 treatment (Figure 3C and 3F). The fold change in Tat-Flag after MG132 treatment reflected the sensitivity of Tat to the proteasomal-dependent degradation pathway. Specifically, more stable Tat corresponded to a lower fold change after proteasome inhibitor treatment. Our results indicated that the degree of fold change was consistent with the stability of Tat. Therefore, the proteasome-dependent Tat degradation pathway was influenced by the carboxyl-terminal region of Tat.
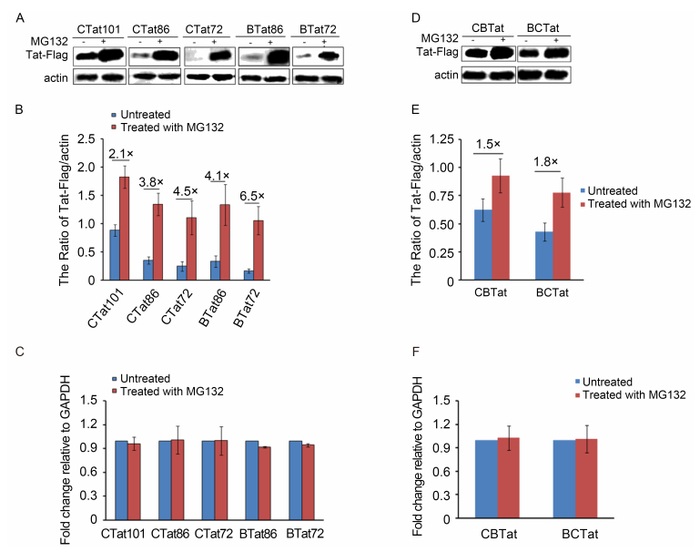
Figure 3. The proteasome-dependent Tat degradation pathway was influenced by the carboxyl-terminal region of Tat. (A, D) HEK293T cells were transfected with Tat-Flag, and treated with (+) or without (−) MG132. Cell lysates were immunoblotted with anti-Flag or anti-β-actin antibodies. (B, E) Quantification of the results in Figure 3A and 3D. Bars represent the relative ratios of Tat-Flag with respect to β-actin levels. (C, F) Quantitative real-time PCR analysis of gene expression in HEK293T cells transfected with Tat-Flag and treated with (+) or without (−) MG132. GAPDH was used for normalization. Two-tailed Student's t-tests were used to assess differences. Means and standard deviations were derived from three independent biological replicates.
-
Transactivation activity differs between full-length B-Tat and the truncated variant Tat86 (Campbell et al., 2005). We thus systematically analyzed the effects of the carboxyl-terminal regions of B-Tat and C-Tat on transactivation activity. We overexpressed Tat-Flag in 293T cells co-transfected with a Photinus luciferase reporter plasmid containing the LTR, i.e., the HIV-1 promoter. When normalized for constitutive Renilla luciferase expression using a plasmid carrying the SV40 promoter, we observed that C-Tat was a significantly stronger transactivator of LTR expression (Figure 4A), consistent with the results of Roof et al. (2002). Overall, B-Tat, C-Tat, and their truncated mutants were all able to promote the transactivation of LTR, but CTat101 was slightly more efficient with respect to LTR transactivation than CTat86 and substantially more efficient than CTat72. Moreover, BTat86 was also slightly more efficient with respect to LTR transactivation than BTat72. These results implied that the carboxyl-terminal region may regulate Tat-mediated transactivation of the LTR region by affecting stability.
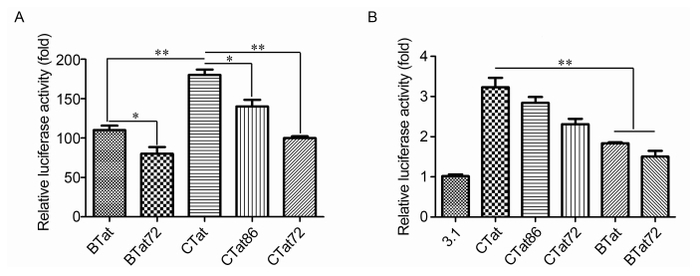
Figure 4. Transcriptional activity regulated by the carboxyl-terminal region of Tat. (A) The effect of truncated forms of Tat on transactivity. HEK293T cells were co-transfected with the pLTR-luc transcriptional reporter plasmid and pcDNA3.1-Tat-Flag plasmid. After transfection for 48 h, luciferase activity was measured. (B) NF-κB-dependent transactivation activity was measured by transient transfection of the 3κB-LUC plasmid and pcDNA3.1-Tat-Flag plasmid into HEK293T cells. After transfection for 48 h, luciferase activity was measured. All samples were co-transfected with a SV40-promoter-containing Renilla luciferase plasmid as a control for normalizing transfection efficiency. * P ≤ 0.05, ** P ≤ 0.01.
To determine the effects of B-Tat and C-Tat on NF-κB activation, luciferase reporter assays were performed using a dual-luciferase reporter assay system by transient transfection of the vector 3κB-LUC and pRL-TK in HEK293T cells. As shown in Figure 4B, C-Tat activated NF-κB more efficiently than B-Tat and the truncated mutants of Tat had a slight effect on Tat-mediated NF-κB activation. In addition, we examined the effect of the carboxyl-terminal region of Tat on transcriptional activity in Jurkat cells, and observed similar results to those using HEK293T cells (data not shown).
The different stabilities of NL4-3 Tat and 1084i Tat
The carboxyl-terminal region of Tat regulates stability
The proteasome-dependent Tat degradation pathway was influenced by the carboxyl-terminal region of Tat
Transcriptional activity regulated by the carboxyl-terminal region of Tat
-
In this study, we found that NL4-3 Tat and 1084i Tat showed different stabilities. Based on an analysis of truncated forms of Tat, we demonstrated that the carboxyl-terminal region regulates stability. We found that 1084i Tat (subtype C) was more stable than NL4-3 Tat (subtype B), consistent with the results of Desfosses et al. (2005). Therefore, it is possible that subtype C is more stable than subtype B.
It has been reported that C-Tat is a significantly stro-nger transactivator of LTR expression than B-Tat (Desfosses et al., 2005). The mechanism underlying differences in transactivation ability between clades B and C Tat is not clear. The N-terminal, cysteine-rich, and core regions of Tat form the so-called transactivation domain, which is primarily responsible for P-TEFb binding. The neighboring basic domain (residues 49-57 in HIV-1 Tat) is essential for regulating TAR RNA binding. Thus, the carboxyl-terminal region of Tat is not directly involved in the Tat-mediated transactivation of the LTR. Figure 4A shows that the Tat-mediated transactivation of the LTR was regulated by the carboxyl-terminal region of Tat. Thus, it is possible that the differences in transactivation between Tat truncated forms result from differences in stability according to the carboxyl-terminal region. In view of the amino acid variation among clade-specific Tat proteins, there may be multiple mechanisms underlying the differences in transactivation between B-Tat and C-Tat. According to our results, the difference in stability between B-Tat and C-Tat may be one explanation.
HIV-1 Tat activates NF-κB by hijacking the inhibitor IκB-α and preventing repressor binding to the NF-κB complex, and the basic domain of Tat is required for the IκB-α association (Fiume et al., 2012). In this study, it was surprising that C-Tat was a stronger activator of NF-κB than B-Tat. As the carboxyl-terminal region of Tat had a slight effect on Tat-mediated NF-κB activation, we compared the basic domains of B-Tat and C-Tat and found a difference at the 57th amino acid. The 57th arginine and serine are highly conserved within both B-Tat and C-Tat (data not shown). Therefore, we hypothesized that the difference at the 57th amino acid within the basic domain between B-Tat and C-Tat may explain the different interactions of IκB-α with B-Tat and C-Tat, which contributes to the differences in NF-κB activation (we are currently examining this hypothesis).
In fact, the clade C virus appears to be transmitted more efficiently than subtype B, as evidenced by its predominance in the HIV-1 pandemic worldwide. In addition, clinical follow-up studies have revealed that subtype C-infected individuals might develop AIDS more easily than those infected with other subtypes (Kanki et al., 1999). This appears to be related to biological differences between viruses, including rapid and high replication rates as well as more efficient transmission rates. Secreted Tat contributes to the spread of HIV-1 via its effect on the surface expression of CCR5 and CXCR4 (Howcroft et al., 1993). Higher Tat stability may contribute to viral transcription and the surface expression of CCR5 and CXCR4. In addition, NF-κB is a master regulator of pro-inflammatory genes and is upregulated in human immunodeficiency virus 1 infection. The mechanisms underlying NF-κB deregulation by HIV-1 are relevant for immune dysfunction in AIDS (Fiume et al., 2012). Therefore, the unique properties of clade C may contribute to its efficient transmission.
-
This work was supported by grants from the National Natural Science Foundation of China (No. 81571987 and 81371820) and the Ph.D. Candidate Research Innovation Fund of Nankai University (2015) (No. 68150003). We greatly appreciate the gift of infectious molecular clone HIV1084i provided by Dr. Charles Wood (University of Nebraska, Lincoln, USA).
-
The authors declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by any of the authors.
-
XCZ and XHK designed the experiments. XCZ, LYQ, DYZ, and DQ carried out the experiments. XCZ and CL analyzed the data. XCZ and XHK wrote the paper. All authors read and approved the final manuscript.

 DownLoad:
DownLoad: