










-
Dear Editor,
Hepatitis C virus (HCV) is a positive-strand RNA virus that belongs to the genus Hepacivirus within the Flaviviridae family. HCV causes chronic liver diseases, and 185 million people are infected (Messina et al., 2015). Currently, there is no approved vaccine to prevent hepatitis C. HCV induces autophagy through elevating reactive oxygen species (ROS) levels via the unfolded protein response (UPR) or via direct interference with the autophagic pathway, which is crucial for the initiation of the HCV infection process (Dreux et al., 2009; Estrabaud et al., 2011; Huang et al., 2013). Autophagy is a highly conserved intracellular process that targets cytosolic components for lysosomal degradation and is initiated by the formation of double-membrane vesicles called autophagosomes (Sica et al., 2015). The auto phagosome ultimately fuses with a lysosome to form an autolysosome (Feng et al., 2014).
The retromer complex is a key part of the endosomal protein sorting machinery that transports cargo proteins to their appropriate destination (Burd and Cullen, 2014). Retromer contains two subcomplexes: a cargo-sorting subcomplex and a membrane-deforming subcomplex. The cargo-sorting subcomplex is a trimer of Vps26, Vps29 and Vps35, whereas an SNX heterodimer forms the membrane-deforming subcomplex (Seaman, 2012). Recently, we discovered that the retromer component Vps35 associates with the HCV NS5A protein and that retromer is required for HCV replication (Yin et al., 2015). In the present study, we found that retromer localized to autophagosomes during HCV replication. Thus, we suggested a link between retromer and autophagy in the context of HCV infection.
To perform the experiments, Huh 7.5.1 cells were grown in Dulbecco’s Modified Eagle’s Medium supplemented with 10% fetal bovine serum. Infectious JFH1 plasmid pJFH1 was obtained from Dr. Takaji Wakita. The OR6 cell line, which harbors full-length genotype 1b HCV RNA and co-expresses Renilla luciferase and from Drs. Nobuyuki Kato and Masanori Ikeda, was grown in DMEM supplemented with 10% FBS and 500 μg/mL of G418 (Promega, Madison, WI, USA). OR6 cells were cleared of HCV infection by incubating with IFN α (10 IU/mL) for 7 days to generate cured OR6 cells. Chloroquine (CQ) was from Sigma-Aldrich (St. Louis, MO, USA).
To study the location of retromer and autophagosome membrane, we perform the immunofluorescence microscopy assay and live cell imaging assay. In the immunofluorescence microscopy assay, Cells seeded onto glass coverslips were washed with phosphate-buffered saline (PBS) and fixed with 4% formaldehyde in PBS buffer for 5 min at room temperature. Fixed cells were stained as previously described (Li et al., 2014). The antibody against LC3B used for immunofluorescence and immunoblot was from Cell Signaling Technology (Danvers, MA, USA) (catalogue No. 3868).
To quantify the degree of colocalization, Pearson correlation coefficient values were calculated using the WCIF Image J software with the colocalization threshold plug-in. Pearson correlation coefficient data was from 32 OR6 cells or 27 JFH1-infected Huh 7.5.1 cells. For Live cell imaging assay, Huh7.5.1 cells grown to 40 to 50% confluency on a chambered coverglass were transfected with GFP-LC3 and mCherry-SNX1 for 24 hours. Time-lapse fluorescence images were acquired with the Ultraview Live Cell Imaging System (PerkinElmer, Waltham, MA, USA). The GFP-LC3 was provided by Dr. Zhiping Xie (Nankai University, Tianjin, China). The mCherry-SNX1 was cloned to mCherry-C1.
Previous work has suggested that HCV uses the autophagosome membrane for RNA replication (Sir et al., 2012). To investigate the role of retromer in HCV induced autophagy, we examined LC3B, Vps35 and NS3 in the context of HCV infection. LC3B localized to membrane only if the autophagy is started. There is no LC3B signal without HCV in cured OR6 cells (Figure 1A). As shown in Figure 1B and 1C, retromer co-localized with autophagosomal puncta in OR6 (Pearson correlation coefficient of LC3B/Vps35: 0.5624 ± 0.0989, n = 32) or JFH1-infected Huh7.5.1 cells (Pearson correlation coefficient: 0.8450 ± 0.0663, n = 27). To determine whether retromer associated with autophagosomes, we performed live cell imaging experiments to assess the localization of GFP-LC3 (an autophagosome marker) and mCherry-SNX1 (a component of retromer) in Huh7.5.1 cells. As shown in Figure 1D, LC3 and SNX1 could localize to the same vesicles. Taken together, these results suggested an association between retromer and autophagosome.
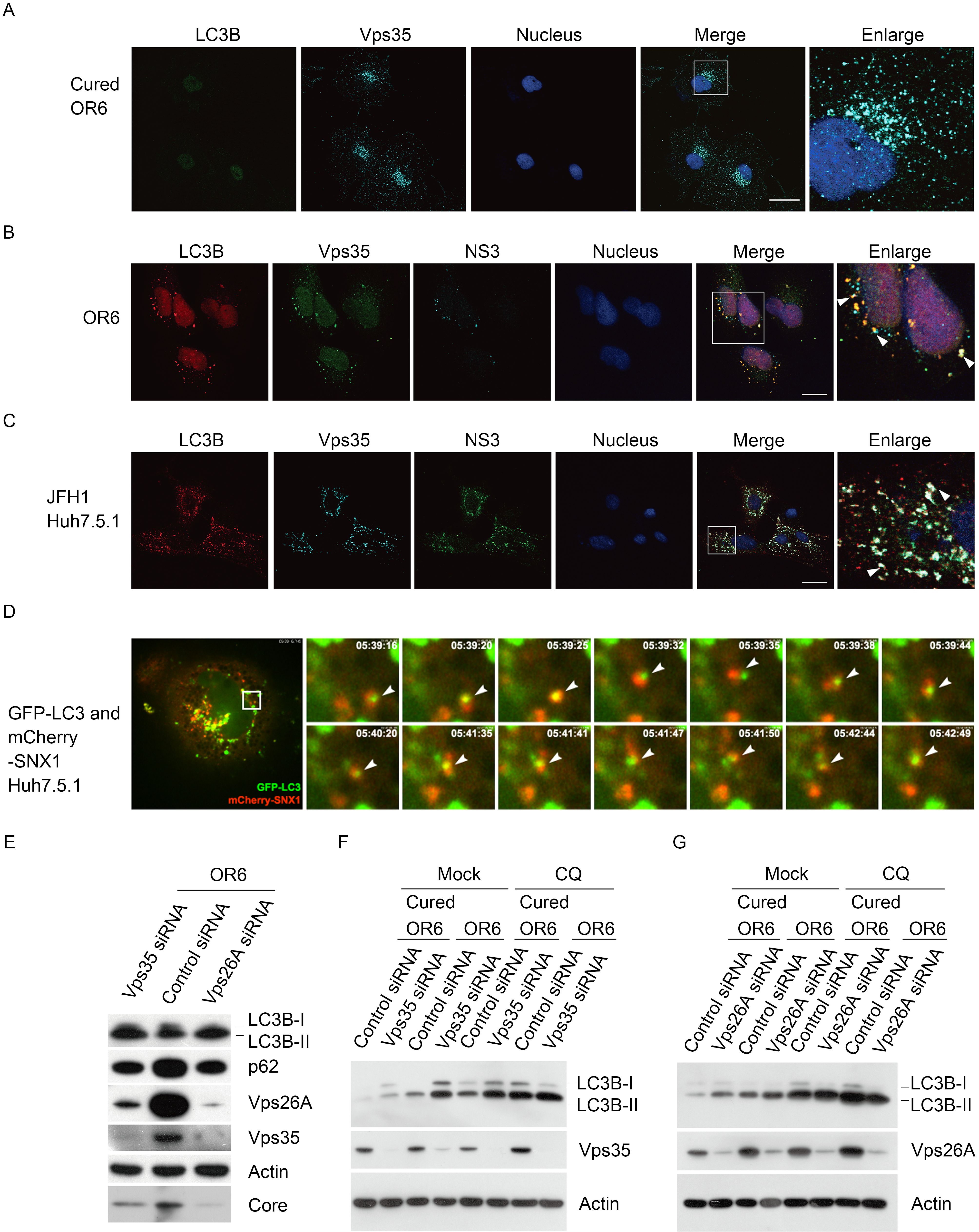
Figure 1. Retromer localizes to autophagosome and is involved in autophagy. (A) Cured OR6 cells were fixed and immunolabeled with anti-LC3B and anti-Vps35. DAPI marks the nucleus. The scale bars represented 10 μm. (B) OR6 cells were fixed and immunolabeled with anti-LC3B, anti-Vps35 and anti-NS3. DAPI marks the nucleus. The scale bars represented 10 μm. (C) JFH1-infected Huh 7.5.1 cells were fixed and immunolabeled with anti-LC3B, anti-Vps35 and antiNS3. DAPI marked the nucleus. The scale bars represent 10 μm. (D) Representative static image at various time points from live cell imaging of Huh 7.5.1 cells transfected with GFP-LC3 and mCherry-SNX1. An example of a vesicle is indicated by the white arrowhead. Time after the start of imaging is presented at the top right corner of each panel. (E) Lysates from OR6 cells treated with siRNA against Vps35 (GCCUUCAGAGGAUGUUGUAUCUUUA), Vps26A (CCACGUAUCCUGAUCUUAA) or control for 72 hours were collected. The samples were immunoblotted for LC3B, Vps35, Vps26A, p62, core, and actin as indicated. (F, G) Cell lysates from cured OR6 or OR6 cells treated with siRNA against Vps35, Vps26A or control for 66 hours and then treated with 50 μmol/L CQ or mock treatment for 6 hours were collected. The samples were immunoblotted for LC3B, Vps35, Vps26A and actin as indicated
To investigate the importance of retromer-autophagosome interaction in HCV replication, we measured protein levels of HCV core, and autophagy markers including LC3B and p62. HCV core protein level was reduced when we knocked down retromer components by siRNA against Vps35 or Vps26A, suggesting that retromer-autophagosome interaction is important for HCV replication (Figure 1E).
To further understand the relationship between retromer and autophagy, we measured autophagic flux by inferring the LC3B-II turnover in the presence and absence of the lysosomal inhibitor chloroquine (CQ), which inhibits autophagy after autophagosomes have formed (Feng et al., 2014). As shown in Figure 1F and 1G, in cured OR6, silencing Vps35 or Vps26A upregulated the level of LC3B-II compared with control siRNA treatment, indicating increased host autophagy. However, in the presence of CQ, the amount of LC3B-II in retromer knockdown or control knockdown OR6 cells is maintained at similar levels, suggesting that the levels of total autophagy including viral induced autophagy and host autophagy remained the same.
Autophagy is commonly exploited as a viral infection strategy (Moser von Filseck et al., 2015). HCV can induce autophagic responses and utilize autophagosomal membranes as sites for the replication of its RNA (Sir et al., 2012). Retromer associates with autophagosomes (Dengjel et al., 2012; Popovic et al., 2012). Interestingly, we noted that retromer colocalized with LC3B in the context of HCV infection, suggesting that retromer may regulate HCV replication through autophagy. We hypothesize that retromer promotes HCV replication by providing autophagosomal membranes. In HCV-infected cells, autophagosomal membranes can be divided into two pools, including host autophagy and viral-induced autophagy. Increased host autophagy by silencing retromer may consume the common membrane resources shared by viral-induced autophagy. As a result, autophagosomal membranes, which support HCV replication, are reduced, and HCV replication is consequently inhibited.
In summary, we propose a model for the role of retromer in HCV replication. Upon HCV infection, retromer may provide double-membrane autophagosomal membranes for HCV replication. Our studies suggested a novel link between retromer and autophagy in HCV replication, which may provide new therapeutic targets for antiviral therapy.
HTML
-
This work was supported by grants from the National Natural Science Foundation of China (81471955, 81672035, 81301438), the CAMS Innovation Fund for Medical Sciences (CIFMS) (2016-I2M-3-020), the National Key Plan for Research and Development of China (2016YFD0500300), the Program for Changjiang Scholars and Innovative Research Team in University (IRT13007), and the Fundamental Research Funds for the Central Universities, the PUMC Youth Fund (3332016085). We thank Guangbo Yang for technical support. We thank Charles Rice, Francis Chisari, Zhiping Xie, Masanori Ikeda, Nobuyuki Kato and Takaji Wakita for reagents. This article does not contain any studies with human or animal subjects performed by any of the authors. The authors declare that there are no conflicts of interest.

 DownLoad:
DownLoad: