











-
Newcastle disease(ND) is a serious avian disease distributed throughout the world that can cause substantial economic losses and remains a major threat to the poultry industry in every country. The aetiological agent,Newcastle disease virus NDV is a member of the genus Avulavirus in the family Paramyxoviridae (11). The genome of NDV is a non-segmented,single -stranded,negative-sense RNA of 15 186,15 192 or 15 198 (2) nucleotides in length,which contains six genes encoding the nucleocapsid protein (NP),phosphoprotein (P),matrix protein (M),fusion protein (F),haemagglutinin-neuraminidase (HN) and large polymerase protein (L). The NP protein binds to the genomic RNA to form a remarkably stable nucleocapsid core structure to which P and L proteins are attached. This ribonucleoprotein (RNP) complex,rather than naked viral RNA,is the template for replication and transcription by the viral polymerase (P and L protein). The NDV genes are linked in tandem in the order 3′-NP-P-M-F-HN-L-5′and are separated by junction sequences that consist of three elements,known as gene-end (GE),intergenic (IG) and gene-start (GS) sequences. For most members of the subfamily Paramyxovirinae,including NDV,efficient replication is dependent on the genome length being a multiple of six. This requirement is known as the 'rule of six' (14).
Reverse genetics manipulation technique is based on intracellular transcription of viral full-length RNAs and simultaneous expression of viral proteins required to form the typical viral RNP complex. The first recovery systems,based on a lentogenic vaccine strain (LaSota) of NDV,were reported simultaneously by two independent groups in 1999 (13, 15). The strategy to recover NDV was to transfect cells with four plasmids encoding the full-length antigenomic viral RNA,the NP protein,and RNA polymerase proteins (P and L) respectively. This transfection resulted in the production of RNPs,leading to the generation of infectious NDV.
Because the rescued virus was wholly derived from cloned cDNA,gene manipulation such as deletion and insertion mutations could be done to investigate virus-host interactions,the molecular basis of NDV pathogenesis,and the strategy for eliminating NDV. Also,using NDV to express foreign gene of other pathogen has been a hot area in vaccine development. Since Krishnamurphy (8) reported that NDV could be used as an expression vector,several genes of other viral protective antigens had been inserted into the NDV genome (6, 12). In this study,we established the reverse genetics system for an NDV ZJI strain of goose origin and inserted the GFP reporter gene between the P and M genes to evaluate the possibi-lity of the expression of a foreign gene in the velogenic NDV and to use the GFP-tagged NDV for investiga-ting the pathogenesis of virulent NDV.
HTML
-
An NDV ZJI strain of goose origin isolated and cha-racterized by Liu et al (5) was grown in embroynated specific pathogen free (SPF) chicken eggs. BSR-T7/5 cells were grown in DMEM containing 10% fetal calf serum
-
Seven cDNA fragments spanning the entire NDV genome were generated by means of reverse transcription PCR (RT-PCR) as described previously (9) and were cloned in pCR2.1 vector (Invitrogen). Using strategically positioned restriction sites,full-length NDV CDNA was assembled in the transcrip-tion vector TVT7R (7) to construct the plasmid pNDV/ZJI.
-
Based on the L gene sequence of NDV ZJI strain,three pairs of primers were employed for PCR and the amplified fragments were joined at the shared restric-tion sites. The resulting fragment was subclon-ed in the eukaryotic expression vector pCIneo-(Promega)between the Xba Ⅰ and Not Ⅰ sites to construct the L expression plasmid pCIneoL.
-
BSRT7/5 cells were grown overnight to 50%~70% confluence in 35-mm dishes. Subsequently the cells were cotransfected with 1 µg full-length cDNA constructs together with 0.5 µg plasmid pCIneoP,1 µg pCIneoNP and 0.5 µg pCIneoL in 15 µL superfect transfection reagent (Invitrogen). After incubation for 3 h,cell cultures were washed and 2 mL fresh medium (DMEM with 10% fetal calf serum) per dish was added. Three days post-transfection,the supernatant and cells were inoculated into the allantoic cavity of 9-day-old embryonated SPF chicken eggs. The inoculated eggs were examined at 12-h intervals and the allantoic fluid was collected following embryo death for virus detection by HA.
-
HA test was done as previously described (4) with allantoic fluid and the positive samples were confirmed by HI test with the monoclonal antibody 6B1 (4) against NDV HN protein.
-
A cDNA fragment containing the GFP gene was obtained from the vector pEGFP (Clontech) and cloned into pCR2.1 vector to construct the plasmid T-GFP with the following primers:
GFP1:5'-ctgggccctcttagaaaaaatacgggtagaagtacc-ATGGTGAGCAAGGGCGAGGAG-3';
GFP2:5'-ttgggccctcTTACTTGTACAGCTCGTC CATG-CCGAGAGTGATCCC-3'
The Apa Ⅰ site is in bold; the NDV gene start and gene end sequences are underlined; the sequence specific to the GFP gene is in uppercase.
The 2 374bp AgeⅠ-PsiⅠ fragment (nt 2 849-5 223) of pNDV/ZJI was amplified and cloned into the pCR2.1 vec-tor with the primers P3(5'-GGGTGAAATGACG-CTCAATAAACTCTC-3') and P4 (5'-ATGGTCTCATC TGTGGCCCGAATACT-3').Subsequently,this frag-ment was subcloned into the plasmid pUC18 resul-ting in pUC AgeⅠ/PsiⅠ. The ApaI site (nt 3134) of this plasmid was used to insert the GFP gene to generate pUC/NDVGFP. The new AgeⅠ-Psi Ⅰ fragment containing the GFP gene was introduced back to full-length cDNA clone after digestion with Age Ⅰ and Psi Ⅰ (Fig. 1). Thus,an additional transcriptional unit,the GFP gene flanked by NDV gene start and gene end signals,was inserted into pNDV/ZJI. The resultant clone was designated pNDV/ZJIGFP. Transfection and recovery of recom-binant GFP-tagged NDV was conducted (Fig. 1) and the recovered recombinant virus was designated as NDV/ZJIGFP.
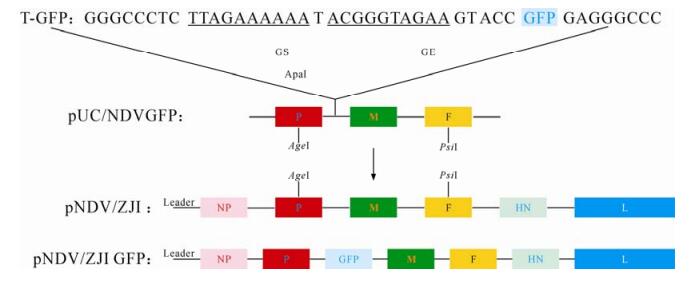
Figure 1. The schematic representation of recombinant plasmid construction
-
Recombinant virus was grown in 9-day-old embryonated SPF chicken eggs. The genomic RNA was extracted from the allantoic fluid using an RNA extract kit (Sangon) and was reverse -transcribed for PCR. With the primers P3 and P4,the Age Ⅰ-Psi Ⅰ fragment containing GFP gene was amplified and cloned into pCR2.1 vector. The positive clone was sequenced to confirm the presence of the GFP gene in the recovered virus.
-
To examine the pathogenicity of the recombinant virus in vivo,the mean death time (MDT) in embryonated chicken eggs and the intrancerebral pathogenicity index (ICPI) test in day-old chickens were performed by standard procedures (1).
-
The recombinant virus was diluted in PBS to give a 10-4 dilution and then was used to infect the BSRT7/5 and CEF cells. The cytopathic effect and the expression of GFP gene were examined at 12h intervals post-infection.
-
Five 10-day-old chickens were inoculated with 104 ELD50 recombinant NDV by the oculonasal route and two chickens were inoculated with sterile PBS as negative controls. Four days post-infection,chickens were euthanized and organs (including brain,lung,spleen,trachea,liver and kidney) were sampled. Due to the instability of GFP during freezing,GFP autofluorescence had to be analyzed in non-fixed tissue samples. Therefore,the segments of tracheae as well as other squeezed thin tissue slices were screened by direct fluorescence microscopy.
1.1. Virus and cells
1.2. Synthesis of cDNA and assembly of a fulllength clone
1.3. Cloning and expression of the L gene
1.4. Transfection and recovery of infectious NDV
1.5. HA and hemagllutination inhibition (HI) test
1.6. Generation of a GFP-tagged recombinant NDV
1.7. RT-PCR and sequencing
1.8. Pathogenicity studies of NDV/ZJIGFP
1.9. Virus growth in cell cultures
1.10. Animal infection test
-
The inoculated embryos were dead 48h post-inoculation and the HA titers were detected as high as 6 log2 similar to the parental virus. The hemagglutination of the rescued virus designated as NDV/ZJI was specifically inhibited by McAb 6B1,proving the successful recovery of authentic NDV/ZJI.
-
After inoculation of transfected supernatant and cells into the embryonated SPF chicken eggs,the recombinant virus was obtained from the allantoic fluid of the dead embryos. The presence of the GFP gene in the genome of recovered virus was verified by RT-PCR with primers P3 and P4. The size of the RT-PCR product from NDV/ZJIGFP was about 3.0 kb while the Age Ⅰ-Psi Ⅰ fragment from NDV/ZJI was 2.3 kb. The sequencing result also demonstrated that the GFP gene was located between the P and M genes in the correct orientation.
-
In order to compare the pathogenicity of NDV/ZJIGFP with the wild-type NDV ZJI strain,MDT and ICPI were examined. The results showed that the MDT and ICPI of NDV/ZJIGFP were 52 h and 1.91 respectively,similar to those obtained for the ZJI strain.
-
Specific green fluorescence was observed in the infected cells 48h post-infection indicating that the GFP gene was expressed at a relatively high level in the recombinant virus; however,CPE could be detected only in the cells infected with NDV/ZJI (Fig. 2).
Figure 2. Infectious test of recombinant virus in cells. A: BSR-T7/5 cell infected with NDV/ZJIGFP; B: BSR-T7/5 cell infected with NDV/ZJI; C: CEF infected with NDV/ZJIGFP; D: CEF infected with NDV/ZJI
-
The fluorescence in the kidneys and tracheae were much more intense (Fig. 3) than that in the other organs (lung,spleen,brain and liver),while no fluorescence could be detected in any organs from chickens inoculated with PBS (data not shown).

Figure 3. Autofluorescence of GFP in the trachea and kidney. A: Fluorescence in the trachea; B: Fluorescence in the kidney
2.1. Recovery of infectious NDV from full-length cDNA
2.2. Generation and identification of infectious recombinant virus
2.3. Pathogenicity studies of NDV/ZJIGFP
2.4. Expression of the GFP gene in infected cells
2.5. Examination of fluorescence in organs
-
In 2005,Liu et al attempted to rescue recombinant NDV/ZJI using support plasmids NP,P and L of ZJI origin without success (10). After replacing three support plasmids of ZJ1 to that of LaSota,NDV ZJI was successfully rescued,but the efficiency of recovery was not high enough for further work. In this study,the full-length cDNA and L expression plasmid were reconstructed,during the process the point mutations accumulated during PCR were mostly minimized to improve the efficiency of recovery. The resultant two plasmids,together with the previous NP and P expression plasmids,were transfected into BSRT7/5 cells to rescue NDV ZJI. Successful rescue was achieved and the experiments were repeated several times with reproducible results,indicating the efficiency of rescue was significantly improved with this newly established system.
Several properties make NDV an attractive vaccine vector for vaccine development against respiratory or intestinal disease in poultry. 1). NDV can be easily cultured to very high titers in embryonated eggs; 2). Mass culture of NDV in embryonated eggs is relatively cheap; 3). NDV vaccines are relatively stable and can be simply administrated by mass application methods such as drinking water or by spraying or aerosol formation; 4). The natural route of infection of NDV is via the respi-ratory and intestinal tract,which are also the major natural routes of infection of other poultry pathogens; 5). NDV can induce local immunity despite of the presence of circulating maternal antibody.
The NDV genes are tandemly linked in the order 3'-NP-P-M-F-HN-L-5'and are separated by junction sequences that consist of three elements,known as gene-end (GE),intergenic (IG) and gene-start (GS) sequences (16).To successfully express foreign gene in the NDV genome,the threeelements need to be maintained such that they flank the target gene. Recently,Park (12) et al. created a bivalent vaccine based on expressing the ectodomain of an H7 avian influenza virus hemaglutinin in a fusogenic and attenuated NDV background. The insertion of the foreign gene (containing only its ectodomain,with the transmembrane and cytoplasmic domains derived from the F protein of NDV) resulted in a chimeric virus with enhanced incorporation of the foreign protein into virus particles.
Because NDV ZJI strain belongs to genotype VIId,which is the predominant genotype in China,the routine vaccines (LaSota and V4) could not provide complete protection against virulent NDV infection. In this study,GFP gene was inserted into the virulent NDV genome and expressed at a high level in the infected cells. Our results demonstrated the possibility of velogenic NDV as a vector to express foreign genes for the first time. Therefore,the ZJI strain can be used to express protective antigens of H5 avian influenza virus,and the vaccine based on the resultant chimera virus will provide better protection against both ND and high pathogenic avian influenza.
Apart from the points mentioned above,the recom-binant GFP-tagged NDV could be also used for inves-tigating the pathogenesis of virulent NDV.GFP-expressing recombinant NDV can be used to monitor infection of cells,embryos or chickens. The advantage of GFP-expressing NDV is that large number of samples can easily be screened for virus replication using direct fluorescence microscopy without any pretreatment. In this research,we demonstrated that strong fluo-rescence could be detected in the tracheae and kidneys,indicating that the trachea and kidney might be chosen as the main target organs for virus isolation. Engel-Herbert (3) et al inserted the GFP gene into the genome of NDV Clone-30,an avirulent strain. Subsequently,the dissemination of NDV was analyzed in the chickens infected with the recombinant virus. Being an avirulent strain,Clone-30 was restricted in the investigation of pathogenesis of virulent NDV. In this regard,the availability of the recombinant virulent NDV (i.e.,NDV/ZJIGFP) would be a useful tool for exploring the pathogenesis of virulent NDV. Work in this respect is now underway in our laboratory.

 DownLoad:
DownLoad: