











-
Severe acute respiratory syndrome virus (SARS-CoV) was the causative agent resulting in the SARS outbreak in 2002-2003 China (5). The identification of SARS-CoV in civet cats and other wild animals in live animal markets revealed that this novel pathogen emerged as a result of an interspecies transmission between animals and hunman (2). In our previous work, we have detected a high sero-prevalence of SARS-CoV infection in horseshoe bats by using a sandwich ELISA kit (Beijing Wantai Biological Pharmacy Enterprise Co., Ltd, China) based on the recombinant nucleocapsid protein (N) of SARS-CoV and an indirect ELISA using inactivated viruses as coated antigen (4). The identification of several different CoVs which are similar to SARS-CoV, called SARS-like CoVs (SL-CoVs), suggests that bats are natural reservoirs of SARS-CoV. According to the phylogenetic analysis, these SARS-and SL-CoVs are grouped into a subgroup of CoV group 2, named group 2b (3, 4, 8).
The SARS-CoV N protein is the most immunogenic protein and stimulates high-level of antibody response in the host and is an ideal viral antigen for detection of virus infection. SL-CoVs are very similar to SARSCoV in genome organization and gene products. In particular, the N proteins share 95%-97% identity (4). Thus the N protein of SARS or SL-CoV can be used as antigen for detection of infections by group 2b viruses.
In this study, SL-CoV N protein was expressed in Escherichia coli, purified and used as antigen. An Indirect Enzyme-Linked Immunosorbent Assay (indirect ELISA) was developed for detection of SARS or SL-CoV infections. This method uses 1-2 μL of serum sample and so can be used for testing of serum sample with limited quantity.
HTML
-
The human cell lines 293T and HeLa were maintained in Dulbecco's modified Eagles medium (DMEM) supplemented with 10% heated-inactivated fetal calf serum (Gibco, USA or Sijiqi, China). HeLa cell lines that stably express human ACE2 protein were established in a previous study (6).
-
Rabbit serum against inactivated SARS-CoV was kindly offered by Dr. Lin-Fa Wang at the Australian Animal Health Laboratory, CSIRO Livestock Industries. AP-conjugated goat anti-rabbit IgG were purchased from PTGLab (Chicago, IL). Horseradish-peroxidase-conjugated protein G was obtained from PIERCE (Rockford, IL).
-
Bat sera were collected from the field as described previously (4).
-
The viral cDNA was prepared from SL-CoV positive bat fecal sample as described previously and used as template (4).
-
Two primers which target the SL-CoV N gene, 5'-CGGATCCATGTCTGATAATGGAC -3' (the enzyme site BamH Ⅰ is underlined) and 5'-GTCGACTTATGCCTGAGTTGAA -3' (the enzyme site Sal Ⅰ is underlined) were used for amplifying the N coding gene. The amplified full-length N gene (encoding 422 aa) was amplified and cloned in frame to the expression vector pET-28b (Novagen) through the BamH Ⅰ and Sal Ⅰ sites fused with six histidine residues. The His6-tagged recombinant N protein was expressed and purified by using a His-bind purification kit (cat No. 70239-3, Novagen) according to the manufacturer's instructions. The concentration of the purified recombinant protein was determined by reading the OD280 value using an Eppendorf biophotometer. Approximately 3 mg of purified protein was obtained from 1 liter of E. coli.
-
Western-blot analysis was performed according to the standard protocol. Briefly, 100 ng of purified fusion protein was loaded into each well of a sodium dodecyl sulphate 12% polyacrylamide gel, then electroblotted onto a nitrocellulose membrane (PIERCE, USA). The membrane was incubated with rabbit serum against inactivated SARS-CoV (diluted at 1:1000), then bound with alkaline phosphatase-labbeld goat anti rabbit IgG. The result was detected by colorimetric assay using NBT/BCIP.
-
The test was modified according to the standard protocol. Each well of the immunoplate (Jincanhua, Shenzhen) was coated with a series of concentrations of the purified (His)6 tagged recombinant N protein overnight at 4℃, then blocked in phosphate-buffered saline (PBS) containing 3% bovine serum albumin (BSA) and 0.05% tween-20 at room temperature for 1 h. After four times washing with washing buffer (PBS containing 0.05% tween-20), 100 μL diluted bat serum (1:100 in PBS containing 3% BSA and 0.05% tween-20) was added to each well of the protein-coated plates, and incubated at 37℃ for 45min. After another four times washing with washing buffer, 100 μL horseradish-peroxidase-conjugated protein G (1:4000 in PBS, 3% BSA, 0.05% tween-20) was added to each well, and the plates were incubated at 37℃ for 45 min. After four washes with washing buffer, 100 μL diluted 3, 3', 5, 5'-tetramethylbenzidine (Sigma, USA) was added to each well and incubated at room temperature for 15 min. The reaction was stopped with 50 μL 0.5mol/L sulphuric acid and the absorbance value was measured at 450 nm. Each sample was tested in duplicate and the mean absorbance value for each sample was calculated. Two rabbit antisera against SARS-CoV and three healthy bat sera were set as positive and negative control respectively.
To establish the baseline for the tests, sera from healthy bats were tested in this assay and the mean absorbance value (Nc) was calculated. The cut-off value was determined as 2.1×Nc. The sample was determined as positive when the absorbance value at 450 was greater than 2.1×Nc.
-
To evaluate the specificity of the indirect ELISA, sera from healthy bat, mouse and rabbit were tested according to the protocol described above. Sera were diluted at 1/100, 1/500 and 1/1000 respectively. The mean absorbance value for each dilution was calculated.
-
Using the protocol described above, we tested for the presence of IgG antibody against the bat SL-CoV N protein in bat serum samples. Rabbit serum against inactivated SARS-CoV and healthy bat serum was used as positive and negative control, respectively.
-
The construction of a codon-optimized full-length spike protein (S) gene of SARS-CoV BJ01 was prepared as described previously (6, 7). Briefly, 12 μg each of pHIV-Luc (pNL.4.3.Luc.E-R-) and plasmid pcDNA 3.1 containing the S gene (or empty vector control) were co-transfected into 2 ×106 293T cells in 10-cm diameter dishes by the standard calcium phosphate method. The pseudoviruses were purified by ultracentrifugation from cell culture supernatant through a 20% sucrose cushion (10 mL) at 55, 000×g for 60 min using a Ty70 rotor (Beckman). The pelleted pseudoviruses were dissolved in 100 μL of PBS and stored at -80℃ in aliquots until they were used. The neutralizing activity of heat-inactivated sera (56℃, 30 min) was determined by mixing 10 ng of pseudovirus (in 30 μL) with diluted antisera (in 30 μL) at 37℃ for 1 h. Sera-pseudovirus complexes were then mixed with 16 ng polybrene (in 40 μL medium) before they were added to human ACE2 expressing HeLa cells. The infected cells were washed with PBS and lysed (Cell Culture Lysis Reagent; Promega) at 48 h post infection. The neutralization activity of each antiserum was monitored by measurement of luciferase intensity as described previously (7). Due to a lack of permissive cell lines for bat SL-CoV, the sera neutralization assay to this virus was not conducted.
Cell lines
Plasmids and antibodies
Bat sera
Viral cDNA
Expression and purification of recombinant SL-CoV N protein
Western-blot analysis
Indirect ELISA
Cross Reaction between the SL-CoV N protein and sera from bat, mouse and rabbit
Seroprevalence of bat SL-CoV infections in bat populations
Pseudovirus-based neutralization assay
-
The N protein was successfully expressed in soluble solution in E. coli and purified. The size of the recombinant N protein was consistent with the expected size of 49.6 kDa (Fig. 1a). Western-blot analysis confirmed that the expressed protein cross reacted with the rabbit serum against SARS-CoV (Fig. 1b).
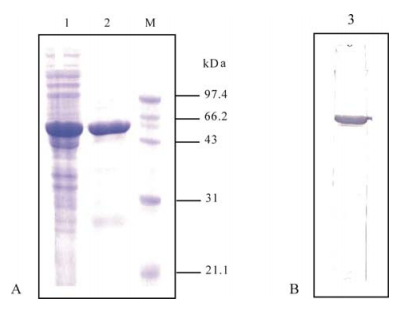
Figure 1. SDS-PAGE (A) and western-blot analysis (B) of the SL-CoV recombinant N protein expressed in E. coli. Lane 1, Supernatant of sonicated cells transformed with pET28b-N; lane 2, Purified recombinant protein; lane 3, Purified recombinant protein probed with serum against SARS-CoV; M, Low molecular protein marker.
-
Different concentrations of the recombinant N protein (1, 2, 10 μg/mL) were used as coated antigen to optimize the amount of coated antigen. The results showed that a concentration of 1 μg/mL is optimal for plate coating (data not shown). The results tested with sera from healthy bat, mouse and rabbit demonstrated that the purified SL-CoV N protein has no cross reactivity when the sera were diluted at 1:100 (data not shown).
-
A total of 573 bat sera, collected during 2005-2008, were tested for presence of SL-CoV infections. As shown in Table 1, most of positive samples were detected in the Rhinolophus species, with a higher prevalence in R. sinicus. These results are inconsistent with previous reports published by ourselves and other teams (3, 4). Furthermore, we have detected SL-CoV infections in Myotis, suggesting that Myotis species are also susceptible to SARS-or SL-CoV.

Table 1. Detection of SL-CoV antibody
The positive sera for the SL-CoV N protein were tested for activity to neutralize the HIV/BJ01. The results showed that none of these positive sera could neutralize SARS-CoV.
Expression and purification of the recombinant SL-CoV N protein
Specificity of indirect ELISA
Seroprevalence of the bat SL-CoV in bats
-
The N protein is a major structure protein in CoV and highly immunogenic in animals. Previous studies have showed that sandwich ELISA based on the recombinant SARS-CoV N protein is sensitive and specific for the serodiagnosis of SARS-and SL-CoV infections (1, 4). However, these methods need 30-50 μL of serum sample and are not capable of testing small quantities of serum sample, e.g. sera collected from small animals like insectivorous bats. In this study we developed an indirect ELISA method using the recombinant SL-CoV N protein as antigen and used it for the detection of SL-CoV antibody. Our results demonstrated that this method is sensitive and specific and can detect SL-CoV infection with 1 μL of bat serum. Further more, the recombinant SL-CoV N protein cross reacted with antisera against SARS-CoV, suggesting that this method can also be used for detection of infections by SARS-CoV. The protein G can bind to the Fc fragment of IgG of many different species. The method developed in this study used can bind to the Fc fragment of IgG of many different species. The method developed in this study used horseradish-peroxidase-conjugated protein G as the secondary antibody and can detect SARS-or SL-CoV infections in human and other mammalian animals.
By using the indirect ELISA developed in this study, we detected 573 bat sera in samples collected from 2005-2008. The results showed that a high sero-prevalence of SL-CoV existed among Rhinolophus species, particularly in R. sinicus, demonstrating that SL-CoVs persistently circulates in Rhinolophus populations, further supporting the proposal that bats are natural reservoirs of SL-CoVs. In addition, we have also detected SL-CoV infections in Myotis, implying that Myotis species are also susceptible to SL-CoV.
It is not surprising that the positive bat sera to SL-CoV N failed to neutralize the protein. The SL-CoVs have a high similarity to the SARS-CoV in most of the gene products, with the exception of the S proteins which share only 76%-78% similarity. This difference in S protein results in different receptor usage by the SL-CoV (7). Thus, antibody detection based on the N protein can be used as a preliminary screen of infections by group 2b CoVs, whereas the sera neutralization based on pseudovirus can be used to differentiate between infections by SARS-or SL-CoV.

 DownLoad:
DownLoad: