









HTML
-
The ability of the Western honey bee, Apis mellifera, to adapt to most climates of the world and the ongoing standardization of colony management has made this species of honey bees the most important species for crop pollination. In recent years, Peru emerged as a main exporter of industrial crops. This industry is mainly concentrated in the Peruvian coastal region, because the local climate permits off-season production of fruits and vegetables that are destined to supply the markets of the Northern Hemisphere. The growth of this agroindustry has directly increased the demand for honey bee colonies for pollination services, thereby promoting local beekeeping. This is in contrast to other parts of the world, where the number of managed colonies and beekeepers has been in general decline over the last decades (Ellis J D, et al., 2010; Potts S G, et al., 2010). The use of pesticides, habitat fragmentation, poor bee management, parasites and diseases have all been pro-posed as possible causal agents for general honey bee colony decline (Oldroyd B P, 2007; vanEngelsdorp D, et al., 2009). Recent analyses of long-term surveys of honey bee colony losses in 16 countries (Neumann P, et al., 2010) pointed to parasites and diseases as the most important causes of general colony losses (Ratnieks F L, et al., 2010).
In Peru, the current status of beekeeping and of honey bee colony health is largely unknown due to the lack of any official institution dedicated to apiculture and due to very few published studies addressing local honey bees. The few previous studies confirmed the admixture of European and African-lineages in Peruvian honey bees due to introgression of African genes into the previously European dominated honey bee populations (Perez-Castro E E, et al., 2002; Quezada-Euan J J G, et al., 2003) as in other Neotropical countries (Smith D R, et al., 1989). Other studies focused on the control of the ectoparasitic mite Varroa destructor (Dávila M, et al., 1987; Tejada G, 1995). However, there are so far no reports about viral diseases in the honey bee populations of Peru. Here we conducted the first survey for viruses in the Africanized honey bees of Peru.
For molecular detection of viruses, adult honey bee workers were collected from 10 queenright colonies at an apiary in the city of Lima on the Peruvian coast. Approximately 100 worker bees from each colony were collected in 96% Ethanol. After transportation, the Ethanol was removed and the samples stored at -20 ℃. Total RNA was extracted from all samples using the NucleoSpin® RNA II kit (Macherey-Nag-el, Dueren, Germany) following the manufacturer’s recommendations. The extracted RNA was eluted in 60 μL of RNAse-free water. BeeDoctor, a MLPA-based diagnostic tool was used for the detection of honey bee viruses (De Smet L, et al., 2012). This RT-MLPA approach detect 6 targets simultaneously covering 10 common honey bee viruses: acute bee paralysis virus (ABPV), black queen cell virus (BQCV), Israeli acute paralysis virus (IAPV), Kashmir bee virus (KBV), deformed wing virus (DWV), Kakugo virus (KV), Varroa destructor virus-1 (VDV-1), slow bee paralysis virus (SBPV), sacbrood virus (SBV) and chronic bee paralysis virus (CBPV). In addition, regular PCR assays were performed for the detection of Lake Sinai virus strain 1 (LSV1) and Lake Sinai virus strain 2 (LSV2). DWV family complex (DWV-KV-VDV-1) and BQCV were detected by BeeDoctor (Figure 1A). No other viruses were detected by regular PCR assays. To test the specificity of the detected viruses, PCR assays were performed with specific primers for the Lp-region of DWV (F1425: CGTCGGCCTATCAAAG; B1806: CTTTTCTAATT-CAACTTCACC) and VDV-1 (F1409: GCCCTGTTCAAGAACATG; B1806: CTTTTCTAATTCAACTTCACC), and for the capsid protein of BQCV (BQCV-F: GTCCAGTGTGATATTGCCAA; BQCV-R: TCATTAGAAAGCG-CCAGACT). For those assays, reverse transcription was performed using M-MLV RT ThermoScript RT-PCR kit (Life Tech-nologies) with 0.5 μg of RNA using random he-xamer oligonucleotides as primers in 20 μL of final reaction volume. The PCR reactions were performed using the MyTaqTM kit (Bioline) with 2 μL of tenfold-diluted cDNA, 2 μL of each primer (10 μmol/L), 10 μL of 5X reaction buffer and 0.25 μL of Taq polymerase (1.25 U) in total 50 μL volume. Positive and negative controls were included for each reaction. PCR amplifications were run in 1.2% agarose gel by electrophoresis, stained in GelRed and visualized under UV light to confirm the PCR product sizes (Figure 1B). The PCR products were purified and sequenced. BLAST runs of those sequences found identities ≥ 97% with DWV and BQCV. These viruses were detected in 70% and 40% of the colonies, respectively. Although our sample size is rather small, those viruses show similar infection rates with a large-scale survey made at same season in a template climatic region (Tentcheva D, et al., 2004). In contrast, virus surveys of Africanized honey bees pe-rformed at a different season in Brazil (Teixeira E W, et al., 2008; Teixeira E W, et al., 2012) showed similar rates for BQCV (37%), lower rates for DWV (20.3%) and the detection of ABPV (27%) and IAPV (25%). These differences in viral rates could be attributed to the season of sample collection and levels of Varroa mite infestation. A mite evaluation of the colonies of the sampled apiary at the beginning of the season found, on average, 3.0 V. destructor mites per 100 worker bees, which is considered to be a low infestation level by local beekeepers. The presence of mites in the colonies may explain the higher occurrence of DWV, a virus vectored by V. destructor, in contrast to BQCV (Ribière M, et al., 2008).
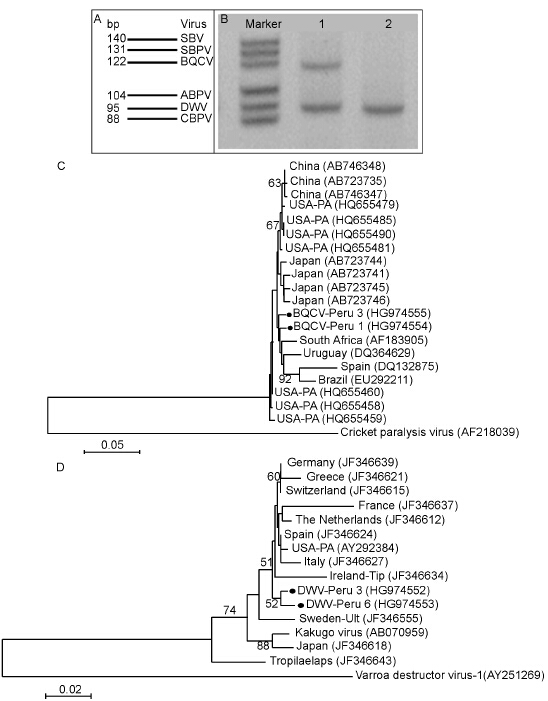
Figure 1. A: Representation of the MLPA markers for six target viruses. Each virus is represented by different band size. B: RT-MLPA amplifications of representative samples. Line 1: Sample positive for BQCV and DWV; Line 2: Sample positive for DWV only. C: Phylogeny of black queen cell virus (BQCV) isolates from Peru (●) and representative isolates from China, USA, Japan, South Africa, Uruguay, Spain and Brazil. Cricket paralysis virus, a related member of the Dicistroviruses, was used for rooting the tree. Nodes supported by bootstrap values >50 are shown. Each isolate is indicated by country of isolation and GenBank accession number. D: Phylogeny of deformed wing virus (DWV) isolates from Peru (●) and representative isolates from honey bees from Europe, USA and Japan; and from Tropilaelaps mercedesae (Acari: Laelapidae) mites from China. Varroa destructor virus-1, a related member of the Iflaviruses, was used for rooting the tree. Nodes supported by bootstrap values >50 are shown. Each isolate is indicated by country of isolation and GenBank accession number.
For the construction of phylogenetic trees appropriate viral isolate sequences from other countries were re-trieved from Genebank. The sequences were aligned using MUSCLE and the phylogeny trees were constructed using the Minimum-Evolution Test bootstrapping se-quences 1000 times with MEGA5 (Tamura K, et al., 2011). The phylogenetic tree for BQCV (Figure 1C) shows, in general, relatively low bootstrap values (< 50) in most of the nodes, but the Peruvian isolates tended to clade together with other South American isolates (Brazil and Uruguay) and with isolates from other continents (Spain and South Africa) and separated from the Japanese, Chinese and US isolates. The phylogenetic tree of DWV (Figure 1D) shows that the Peruvian isolates positioned in a very homogeneous clade composed by the US and most of the European isolates. Although, this clade shows many nodes with relatively low bootstrap values (< 50) it nevertheless indicates that the Peruvian DWV isolates derived from this monophyletic group that is well separated from DWV isolates from A. mellifera and Tropilaelaps mites isolated from Japan and China, respectively.
Finally, the lack of detection of the other ten viruses tested could indicate there is a low frequency of these viruses in the surrounding Africanized honey bee po-pulations, but a more extensive survey will be necessary to draw definitive conclusions.
-
Financial support was provided to OY and PN by the COST funded project VIVA and the Swiss National Science Found-ation (SNF). The funding agencies were not involved in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the paper for publication.
All the authors declare that they have no competing interest. This article does not contain any studies with human participants or animals performed by any of the authors.

 DownLoad:
DownLoad: