










HTML
-
Despite an effective vaccine, hepatitis B virus (HBV) infection remains a major global health problem, affecting 248 million people worldwide (Schweitzer et al., 2015). More than 780, 000 people die annually because of hepatitis B related secondary diseases, primarily cirrhosis and hepatocellular carcinoma (Komatsu, 2014). Antiviral therapies developed during the past 20 years, such as nucleos(t)ide analogs (NA) and interferon-α (IFN-α), are effective in suppressing, but rarely in eliminating, HBV infections (Chen and Yuan, 2014; Hadziyannis, 2014; Liu et al., 2014; Koumbi, 2015). INF-α may help degrade nuclear viral DNA, but the effect is limited and less than 10% of patients show a sustained virological response measured as loss of hepatitis B surface antigen (HBsAg) (Isorce et al., 2015). NAs are effective inhibitors of the HBV reverse transcriptase (RT) enzyme and can prevent the release of infectious virions from HBV infected cells. However, cessation of treatment can result in viral relapse since NAs alone have little or no effect in the elimination of the replicative template of HBV, covalently closed circular DNA (cccDNA) (Lucifora et al., 2014; Yang and Kao, 2014). Consequently, in order to effectively clear HBV infection and produce full remission, there is an urgent need for a different treatment paradigm to inhibit HBV replication and eliminate latent viral reservoir cccDNA. Type II bacterial clustered regularly interspaced palindromic repeat (CRISPR)-associated (Cas) 9-based genome editing technologies provide a potential solution (Perkel, 2015).
Genome editing is a novel approach that enables investigators to manipulate target genes in various cell types and organisms by using engineered nucleases. In the past 3 years, the CRISPR/Cas9 system has been engi neered into an accurate and efficient genome editing tool and exhibits the potential to engineer and modify DNA sequences in diverse species because of its simple design, construction, and application (Fu et al., 2013; Ran et al., 2015). The CRISPR/Cas9 system has been successfully applied to human cells to inhibit invasive pathogens and it promises to be applicable as a therapeutic strategy for HBV infections (Lin et al., 2014; Yin et al., 2014; Ramanan et al., 2015). This review aims to summarize recent advances in the application of CRISPR/Cas9 as an inhibitor of HBV, and discuss the obstacles and possibilities of the CRISPR/Cas9 system as a curative therapy for chronic hepatitis B infection.
-
HBV, a member of the family Hepadnaviridae, is one of the smallest enveloped DNA viruses, with a 3.2 kb-long circular genome (Ganem and Varmus, 1987; Datta et al., 2012; Seeger and Mason, 2015). A remarkable feature of the HBV genome is its extremely compact organization, containing four overlapping open-reading frames (ORFs) C, S, P, X that encode Core proteins (Core and preCore), surface antigen proteins (PreS1, PreS2, and S), reverse transcriptase (Pol protein) and X protein, respectively (Beck and Nassal, 2007; Datta et al., 2012). Hepatitis B surface antigen (HBsAg) envelops the viral nucleocapsid, which is formed by the core protein (HBcAg). The encapsidated viral genome is organized as a relaxed circular partially double-stranded DNA (rcDNA) (Dryden et al., 2006). Upon infection of hepatocytes, the HBV rcDNA is converted by cellular enzymes into cccDNA inside the nuclei of infected cells by a DNA repair mechanism that is still not understood in detail. Episomal HBV cccDNA exists persistently in hepatocyte nuclei as a stable minichromosome organized by histone and non-histone proteins and acts as a viral transcription template, which utilizes the cellular transcriptional machinery to produce all viral RNAs necessary for protein production and viral replication (Nassal, 2015).
HBV cccDNA transcribes four viral RNAs, known as pregenomic RNA (pgRNA), preS, M/S, and X RNAs (Seeger and Mason, 2015). The pgRNA serves as mRNA for the viral proteins or as a template for the viral genomic DNA through reverse transcription. Cytoplasmic pgRNA and the P protein (viral reverse transcriptase) are co-packaged into viral capsid, and rcDNA is produced from the reverse transcription of pgRNA (Figure 1). Nucleocapsids containing rcDNA are released from the host cell as virions or are converted to cccDNA in the nucleus (Werle-Lapostolle et al., 2004). HBV cccDNA exhibits staggering stability and declines slowly under present antiviral therapies. Therefore, controlling and eradicating cccDNA is the critical obstacle to effective treatment of HBV infection (Levrero et al., 2009; Schiffer et al., 2012; Nassal, 2015). Theoretically, specific disruption of the HBV genome to eradicate cccDNA may cure chronic HBV infection completely.

Figure 1. Life cycle of HBV. HBV virions enter the hepatocyte by binding to the receptor NTCP, and possibly other unknown receptors. After uncoating, the core particles are delivered to the nucleus, where the rcDNA is converted into cccDNA. cccDNA persists in the nucleus as a minichromosome that serves as the template for transcription of viral RNA including pgRNA and subgenomic RNAs. The pgRNA in the cytoplasm is translated into the core protein and the viral polymerase; the subgenomic RNAs are translated into envelope proteins and the X protein. The pgRNA is then packaged into capsid particles together with the viral polymerase and is reverse-transcribed into rcDNA. The resulting core particle can either be enveloped with surface antigens in the ER and released from the hepatocyte as progeny virions or be reimported to the nucleus for additional cccDNA amplification. Viral cccDNA is the target of CRISPR/Cas9 gene editing. Abbreviations: cccDNA: covalently closed circular DNA; pgRNA: pregenomic RNA; rcDNA: relaxed-circular DNA; P: HBV polymerase; TDP2: Tyrosyl DNA Phosphodiesterase-2.
-
The CRISPR/Cas system was originally identified as an adaptive RNA-mediated immune system in bacteria, which rejects invading bacteriophages by introducing targeted DNA mutations into pathogenic viruses and plasmids (Haft et al., 2005; Garneau et al., 2010; Horvath and Barrangou, 2010). In this process, CRISPR RNA (crRNA)-guided Cas proteins are employed to recognize target sites within the invader's genome. In 2012, the system was simplified to target any DNA sequence from virtually any organism, with the development of modified CRISPR components comprising a short chimeric single guide RNA (sgRNA) and a Cas9 nuclease from Streptococcus pyogenes (Wang et al., 2013; Yang et al., 2013). Later, two groups successfully edited a mammalian genome using CRISPR/Cas9 (Cong et al., 2013; Mali et al., 2013). In the CRISPR/Cas9 system, sgRNA directs Cas9 DNA endonuclease to the target DNA sequence next to the protospacer adjacent motif (PAM) for site-specific cleavage and produces sequence-specific double-strand breaks (DSBs) (van der Ploeg, 2009). Consequently, various mutations such as substitutions, deletions and insertions in the target genome are introduced by the host DNA repair machinery (Wyman and Kanaar, 2006) including nonhomologous end joining (NHEJ) at the binding site (Matthews and Simmons, 2014), or, alternatively, homologous-dependent repair (HDR) (Smith, 2001; Doudna and Charpentier, 2014; Xu et al., 2014).
Compared with zinc finger nuclease (ZFN) and transcription activator-like effector nuclease (TALEN)-based genome editing, CRISPR/Cas9 can easily be reprogrammed to cleave virtually any DNA sequence by simply designing a single RNA sequence that matches the DNA targeted for cleavage (Choi and Meyerson, 2014; Xu et al., 2014). In addition, because the sgRNA component is physically separate from Cas9 expression, the sgRNA is easily "programmable", with the possibility for many sgRNAs targeting multiple DNA sites when expressed simultaneously with the same Cas9 (Cong et al., 2013). Therefore, CRISPR/Cas9 has shown great promise in realizing potent and multiplex genome editing and regulation of gene expression without host dependence. CRISPR/Cas9-based tools have been successfully applied in diverse organisms and in a broad range of research fields, including high throughput genetic screens (Shalem et al., 2014), generation of gene knockouts in several species (Yin et al., 2014; Arazoe et al., 2015), and targeting of pathogens to eradicate infections such as HIV and HBV (Figure 2) (Ohno et al., 2015; Saayman et al., 2015; Strong et al., 2015; Zhu et al., 2015).
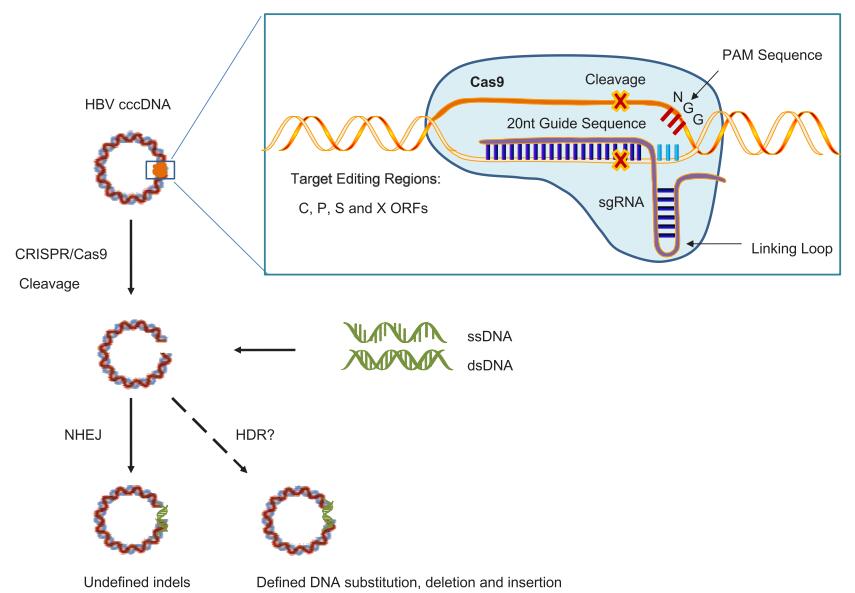
Figure 2. Application of CRISPR/Cas9 in targeted HBV genome editing. CRISPR/Cas9 system consists of a Cas9 nuclease and a single guide RNA (sgRNA), which leads Cas9 to the target site. The sgRNA includes a dual-RNA sequence derived from CRISPR RNA (crRNA) and a separate transcript RNA (tracrRNA) that binds together with a linking loop. The sgRNA guides the Cas9 endonuclease via matching the 20-nucleotide sequence of targeted genomic DNA to produce site-specific double-strand breaks at positions that are 3 bp upstream of protospacer adjacent motif (PAM) sites (indicated by arrow). In published studies, various target sites in the HBV genome, including all four ORFs (C, P, S and X), were selected as targets in designing HBV-specific sgRNA. Cas9/sgRNA-mediated HBV DNA cleavage introduces blunt double-stranded DNA breaks in genomic loci, which become substrates for endogenous cellular DNA repair machinery that catalyzes error-prone nonhomologous end joining (NHEJ) to rejoin the ends and introduces undefined small deletions and additions (indels) to confer viral DNA substitution or deletion. Up to now, no study showed that homologous-dependent repair (HDR) was involved in repairing the HBV cccDNA cleaved by CRISPR/Cas9. Red asterisks: cleavage sites.
-
Specifically targeting HBV DNA enabling deactivation or elimination using the CRISPR/Cas9 system is an attractive goal aimed to cure HBV. Selecting effective and specific target sequences in the viral DNA genome to design the HBV-specific sgRNA is the first and critical step in constructing the CRISPR/Cas9 system. In published studies, various target sites in the HBV genome, including the four ORFs (C, P, S and X), were selected as targets in designing HBV-specific sgRNA. To avoid off-target effects and minimize toxicity, it is necessary to use conserved sequences in the HBV genome and avoid similar parts of the human genome. Based on studies published by September 2015, HBV antigens, its DNA genome and cccDNA were all found to be significantly suppressed by certain HBV-specific gRNAs during in vitro or in vivo experiments (Table 1) (Lin et al., 2014; Seeger and Sohn, 2014; Dong et al., 2015; Karimova et al., 2015; Kennedy et al., 2015a; Liu et al., 2015; Ramanan et al., 2015; Wang et al., 2015; Zhen et al., 2015)

Table 1. Published studies on the application of CRISPR/Cas9 system to inhibit HBV replication.
In detail, it was reported that transfection of Cas9/ sgRNAs into cultured cells decreased HBV DNA release by 77% to 98% with different sgRNAs (Ramanan et al., 2015). The total amount of cccDNA was shown to decrease by 60.6% to 75.0% in Huh7 cells transfected with pCas9 (Dong et al., 2015). Kennedy's group reported that HBV RT-specific sgRNA repressed cccDNA formation by 10-fold, while surface Ag and core-specific sgRNAs sup pressed cccDNA levels by 4-fold (Kennedy et al., 2015a). Lin et al. used a duck hepatitis B virus (DHBV)-expressing plasmid and found that DHBV-specific gRNAs only slightly reduced the levels of the DHBV-expressing plasmid but efficiently suppressed the levels of rcDNA and cccDNA (Lin et al., 2014). In vivo, immunoblots revealed that only a minimal amount of the intracellular core protein was produced in pCas-injected mouse liver. Quantification of intrahepatic cccDNA also showed a reduction on CRISPR/Cas targeting (Dong et al., 2015).
Of note, various genome targeted gRNA/Cas9 systems showed various suppression efficiencies on HBV DNA and antigen expression. For example, surface Ag-specific sgRNA suppressed HBsAg production more effectively than other sgRNAs, with the inhibition rate ranging from 60% to almost 100% (HBsAg undetectable) with different cell culture systems and transfection methods (Lin et al., 2014; Kennedy et al., 2015a). Cells transduced with RT specific sgRNA showed a statistically significant reduction in HBeAg secretion. In contrast, there was no significant reduction in HBeAg for core and surface sgRNAs (Kennedy et al., 2015a). Dong et al. (2015) and Lin et al. (2014) reported that X region specific sgRNA exhibited powerful CRISPR/Cas9-mediated inhibition of HBV, perhaps because X encodes a protein that regulates viral gene transcription and is required for efficient viral replication and spread (Tang et al., 2006). The P region was another efficient site for specific targeting of sgRNA in suppressing HBV DNA replication and antigen transcription. Results from Kennedy et al. showed that RT-specific sgRNA, which targets the essential "YMDD" motif in the HBV P ORF for cleavage, essentially entirely blocked virus replication, shown by a reduction in total viral DNA released into the culture medium of 1000-fold, total intracellular HBV DNA levels decreased by 100-fold, and inhibition of the accumulation of cccDNA by up to 10-fold (Kennedy et al., 2015a).
One advantage of CRISPR/Cas9 compared with other gene editing methods is that it has the capacity for multiplex targeting by providing a method for multiple disruptions, insertions, and deletions with high efficiency and low cost. Therefore, the combination of different specific site sgRNAs targeting several sequences in the HBV genome for multiple genomic editing will be an ideal way to improve the suppression efficiency. Lin et al. reported that the combination of sgRNAs P1 and XCp was more effective in suppressing intracellular HBsAg production than either sgRNA alone (Lin et al., 2014). In vivo, the in-hibitory effect of CRISPR/Cas9 on serum HBsAg levels was highest (93%) for a combination of X and S specific sgRNAs (Zhen et al., 2015). However, combination of multiple sgRNAs also produced side-effects in the results of Wang et al., who found that the specific DNA fragment between the two cleavage sites of the gRNAs was removed by use of the dual gRNAs (Wang et al., 2015). Based on the fact that specific sgRNAs and combinations may result in differences in editing efficiency, strategies for specific HBV region sgRNA design and combination should be precisely verified in future studies.
-
In addition to cleavage, functional inactivation of HBV DNA (in particular cccDNA) caused by mutation also contributes to the inhibitory effects of CRISPR/Cas9 in suppressing HBV replication. Cas9 cleavage of targets in the residual viral DNA usually results in the introduction of small sequence insertions or deletions (indels), which can be assessed by T7 endonuclease (T7E1) assay and DNA sequencing. Dong et al. reported that an amplified PCR fragment of cccDNA was cleaved into 240 and 540 bp pieces in pCas9-1-transfected cells and into 380 and 400 bp pieces in pCas9-2-transfected Huh7 cells co-transfected with HBV precursor plasmid precccDNA, judged by T7E1 assay (Dong et al., 2015). Another study in Huh7 cells transfected with vector pAAV/HBV1.2 reported that the mutation rate in HBV expression templates edited by sgRNAs was 9.3%-13.6% with single sgRNAs, and up to 25.6% with a combination of sgRNAs (Lin et al., 2014). When sequencing DNA fragments amplified in the T7E1 assay, 58%–75% of the amplified HBV sequences contained mutations in S-and X-sgRNA/hCas9-treated mice (Zhen et al., 2015). The most frequent mutations (66%) were single-nucleotide deletions or insertions, followed by deletions that spanned > 100 nucleotides (19%), then deletions of < 30 nucleotides (12%) (Seeger and Sohn, 2014). These mutation patterns of HBV induced by CRISPR/Cas9 are observed as a result of DNA repair in the non-homologous end joining (NHEJ) pathway and typically occurred after genome editing(Cong et al., 2013; Seeger and Sohn, 2014; Dong et al., 2015; Zhen et al., 2015). In contrast, up to now no study showed that HDR was involved in repairing the HBV cccDNA cleaved by CRISPR/Cas9. However, with the introduction of homologous DNA sequence, HDR can be expected as a method in CRISPR/Cas9 system to specifically define HBV DNA at planed sites, such as sequence substitution, deletion and insertion (Figure 2).
Interestingly, although the mutation rates of the HBV genome are modest in vivo, the decrease in HBV DNA is quite remarkable in these studies. This inconsistency of mutation and clearance rates of viral templates might be attributed to fragments of HBV template cleaved by sgRNA/Cas9 not being completely rejoined by NHEJ; most of them may be cleared by other undefined mechanisms (Liu et al., 2015).
-
Current therapeutic treatments for chronic HBV infection include the use of INF-α and NAs, such as lamivudine (3TC), tenofovir disoproxil fumarate (TDF) and entecavir (ETV) (Koumbi, 2015). The fact that they cannot eliminate HBV replication completely has prompted the search for combination therapies using CRISPR/Cas9 with NAs or IFN-α, aimed at eliminating latent viral reservoirs. Experimental results show that IFN-α neither has a measurable effect on the antiviral activity of the CRISPR/ Cas9 system, nor affects the mutations of cccDNA (Seeger and Sohn, 2014). In contrast, treatment of HepG2.2.15 cells with HBV specific sgRNA transfection as well as TDF, ETV, or 3TC demonstrated a clear effect, at least additive, leading to more efficient elimination of residual HBV DNA replication. Significantly, the residual HBV DNA after the use of ETV was almost completely inhibited on application of the CRISPR/Cas9 system, which further confirms the potential of this approach to very efficiently edit cccDNA (Kennedy et al., 2015a). Thus, CRISPR/Cas9 might complement the presently available HBV antiviral treatment regimens, especially NAs, providing therapeutic approaches against targets that include HBV latent viral reservoirs of cccDNA.
Various HBV-specific sgRNA design regions and suppression efficiencies
CRISPR/Cas9-mediated mutation of the HBV genome
Combined application of CRISPR/Cas9 with other antiviral treatment
-
Although the progress of research on the application of the sgRNA/Cas9 system in suppressing HBV has been impressive, several important obstacles remain (Kennedy et al., 2015b; Koskella, 2015). Much more research is required to enable practical use and there is ample room for other CRISPR/Cas9 strategies to improve efficiency and specificity in inactivating and eradicating HBV (Kennedy and Cullen, 2015).
-
To achieve continuous and prolonged treatment in vivo, an efficient delivery system for CRISPR/Cas9 would be used to take the targeting nucleases precisely to infected cells. Efficient access of the nucleases would be required to all cccDNA molecules to eradicate HBV. In the current studies, Cas9/sgRNAs transfected by lentivirus vector induced robust suppression of secreted HBsAg, normally over 90%, indicating lentiviral vectors enable the constitutive expression of Cas9 and/or sgRNAs in vivo (Dong et al., 2015; Zhen et al., 2015). However, lentiviral vectors cannot be safely used in the clinical setting because they can induce immune responses and can be integrated to the host genome causing insertional mutagenesis (Annoni et al., 2013). Adeno-associated virus (AAV) vectors can be generated at very high titers with a high degree of tropism for hepatocytes, so are promising viral vectors to efficiently deliver the designed CRISPR/Cas9 into the liver for anti-HBV therapy (Asokan et al., 2012). However, the currently used S. pyogenes Cas9 gene, at 4.2 kb, is too large for incorporation into AAV vectors, as these can only accommodate 4.6 kb of DNA, which must include not only the Cas9 ORF but also the sgRNA and the transcriptional regulatory regions required for Cas9 mRNA and sgRNA expression. To solve this problem, Cas9 genes from other bacterial species can be considered. Several smaller Cas9 genes have been found and two of them, expressed by Neisseria meningitidis (Esvelt et al., 2013) and Staphylococcus aureus (Ran et al., 2015), have been characterized and raise the possibility of generating AAV vectors able to deliver Cas9/sgRNA combinations specific for HBV to the liver in vivo with high efficiency (Kennedy et al., 2015a). However, this approach needs more experiments for optimization and to prove the efficiency.
-
Off-target effects are pertinent concerns for all genome-editing strategies including the CRISPR/Cas9 system, because they may lead to non-specific gene modification events such as chromosomal integration of the linearized viral DNA, and consequently adversely affect target cell function, especially by the long-term presence of the effector nucleases. Although low-to-absent levels of off-target effects have been detected in Cas9-directed HBV genome cleavage within the genome at sites of close homology to the intended target, the possibility cannot be ignored because in previous study off-target effects were observed with up to five mismatches of the host chromosomal DNA sequence to the sgRNA (Fu et al., 2013). Moreover, DNA repair after the cleavage of cccDNA can induce mutations of the HBV genome and some specific mutations may be associated with accelerating carcinogenesis in the host (Park et al., 2014). To decrease the danger of these non-specific effects, significant effort has been made, including truncating sgRNAs (Fu et al., 2014), generating paired nickase sgRNAs (Ran et al., 2013; Shen et al., 2014; Gebbing et al., 2015) and dCas9-FokI obligate heterodimers (Tsai et al., 2014), and the use of protein-destabilizing domains (DD) (Iwamoto et al., 2010; Sellmyer et al., 2012). DD-Cas9 fusions would allow for temporal and dose-dependent control of Cas9 activity by degrading the fusion protein with a DD-specific small molecule inhibitor. This inducible expression system can thereby be stringently controlled and has the potential to avoid potential off-target effects from constitutive Cas9 activity by modulating the concentration and the timing of the treatment intervention (Saayman et al., 2015).
-
Most studies to date used designed HBV-specific sgRNA against the HBV ORFs (C, P, S and X). However, HBV replication is a complex process involving both viral and host factors. Therefore, it may be possible to identify additional sgRNA targets in the HBV genome or host that might inactivate HBV, including cccDNA, with a significantly higher efficiency and specificity than in the reports so far. These target sites might include upstream promoter sequences in the HBV genome, potential conserved viral and host lncRNAs that could modulate HBV replication, host dependency factors such as HBV receptor-sodium taurocholate co-transporting polypeptide (NTCP) (Yan et al., 2013; Yan et al., 2014), or histone and non-histone proteins involved in stable cccDNA minichromosome organization. Moreover, because integration of HBV DNA fragments into host genome might promote hepatocellular carcinoma(HCC) development, future efforts can also be considered to focus on developing multiple gRNAs or combinations to remove integrated HBV DNA from the host genome to cure HBV-related HCC (Hai et al., 2014; Wang et al., 2015).
Improve efficiency of the delivery system
Avoidance of off-target effects
Optimization of target sites
-
In recent years, CRISPR/Cas9 technology has been regarded as a cornerstone of bioscience for its potent power to edit genomes and control gene expression. Experimental studies have shown that CRISPR/Cas9 disrupts episomal cccDNA, reduces pregenomic RNA, and suppresses viral proteins both in vitro and in vivo without any cytotoxic effects, providing substantial proof for the idea that the CRISPR/Cas9 system is a promising technology with the potential to function as an effective treatment for HBV infections, and, in particular, to eradicate cccDNA. However, it is important to be cautious about the challenges that lie ahead as the feasibility and safety of these novel approaches remain uncertain in practical use. Numerous issues must be resolved and there is still plenty of room for improvement to achieve safer and more effective therapeutic options to use CRISPR/Cas9 genome editing technology for sustained HBV inhibition and cccDNA eradication.
-
This work was supported by National Natural Science Foundation of China (NSFC8120232, NSFC81461130019), Transregio-SFB (TRR) of the Deutsche Forschungsgemeinschaft (DFG TRR60).
-
The authors declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: