









-
Dear Editor,
Due to its high transcription efficiency, bacteriophage T7 RNA polymerase (T7 RNAP) has long been utilized in bacterial and in vitro systems to generate large quantity of RNA for various purposes (Studier and Moffatt, 1986). However, heterogeneity at the 3′-end of the RNA transcript remains a major limitation for in vitro RNA preparation using T7 RNAP. When approaching the end of its DNA template, T7 RNAP tends to add a few extra nucleotides not directed by the template strand sequence, resulting in a mixture of RNA products that are equal to or longer than the desired length (Milligan et al., 1987). Various strategies have been used to possibly overcome this limitation, such as introducing 2′-methoxy groups to the last two nucleotides at the 5′-terminus of the DNA template strand (Kao et al., 1999), seeking other high-efficiency RNA polymerase (Zhu et al., 2014), using a general purpose RNA-cleaving DNA enzyme (Santoro and Joyce, 1997), or introducing a self-cleaving ribozyme at the 3′-end of the target RNA sequence (Kieft and Batey, 2004). We previously utilized the glmS (a bacteria gene that encodes a glucosamine-6-phosphate synthase) ribozyme approach to generate homogenous RNA samples for crystallographic and biochemical characterizations of picornavirus and flavivirus RNA-dependent RNA polymerases (RdRPs) (Gong and Peersen, 2010; Wu et al., 2015). The glmS ribozyme contains an “AG” sequence at its 5′-end and the self-cleavage site is between these two nucleotides. As a consequence, the target RNA after glmS self-cleavage must bear an “A” at its 3′-end, limiting the designing space of the target RNA. When a non-adenosine nucleotide was required at the RNA 3′-end, we took the chemical synthesis as the solution. For example, a chemically synthesized T30 RNA construct bearing a cytosine at the 3′-end (Integrated DNA Technologies, Table 1 and Figure 1A) was used in a dinucleotide-driven de novo RdRP assay characterizing the Japanese encephalitis virus (JEV) NS5 RdRP (Wu et al., 2015). In this study, we have attempted to practically resolve the aforementioned limitation of the glmS approach by replacing the glmS ribozyme with the Hepatitis delta virus (HDV) ribozyme. The HDV ribozyme is the only known ribozyme with almost no prerequisite on 5′-sequence of the cleavage site (Perrotta and Been, 1992). Our data demonstrate that the HDV ribozyme-derived approach is capable of generating the T30 construct. After post-cleavage treatment to remove the 2′, 3′-cyclic phosphate of its 3′-terminal nucleotide, it is able to drive de novo RNA synthesis of the West Nile virus (WNV) NS5 RdRP as efficiently as the chemically synthesized RNA construct.
Name Sequences (5′–3′) Length (nt) T30 GGGAGAUGAAAGUCUCCAACGAUUAUAUCC 30 primer A GAATTC TAATACGACTCACTATA GGGAGATGAAAGTCTCCAACGATTATATCC GGGTCGGCATGGCATCTCCACC 75 primer B GCCGGC CCTTCTCCCTTAGCCTACGAAGTAGCCCAGGTCGGACCGCGAGGAGGTGGAGATGCCATGCCGACCC GG 75 5′-GEN GCGCGCGAATTC TAATACGACTCACTATAG 30 3′-HDV ACCGGTGCCGGC CTTCTCCCTTAG 24 Note: Restriction enzymatic sites are shown in italic, and the 24-nt shadowed sequences in primers A and B are complementary and are designed for the primer extension reaction. The singly underlined, bold, and doubly underlined sequences correspond to the T7 RNAP promoter, T30, and part of the HDV67 region, respectively. Table 1. Nucleotide sequences of the de novo synthesis RNA template T30 and the oligonucleotides used in the construction of the pRAV23-T30-HDV67 plasmid
For construction convenience, a modified 67-nculeotitde (nt) HDV ribozyme sequence (HDV67, Figure 1A) containing a shortened P4 region (P4 stands for “base-paired region 4”) was chosen for the ribozyme replacement cloning process (Schurer et al., 2002). The pRAV23 plasmid (kindly provided by Dr. Jeffrey Kieft, University of Colorado Denver, USA and Dr. Robert Batey, University of Colorado Boulder, USA) was used as the template. First, two DNA constructs primer A and primer B (Table 1) were used in primer-extension reactions to generate double-stranded DNA including both the T30 and HDV67 sequences. Polymerase chain reaction (PCR) was carried out to amplify this DNA, and the PCR primers 5′-GEN and 3′-HDV (Table 1) also introduced EcoR I and NgoM IV restriction sites at the product termini, respectively. Subsequently, restriction enzyme digestion was carried out for both the PCR product and the pRAV23 plasmid, and the desired fragments (gel-purified if applicable) were then ligated using bacteriophage T4 DNA ligase to generate the pRAV23-T30-HDV67 plasmid. The resulting plasmid was then transformed into Escherichia coli (E.coli) DH5α competent cells for plasmid preparation, and sequence of interest was confirmed by sequencing.
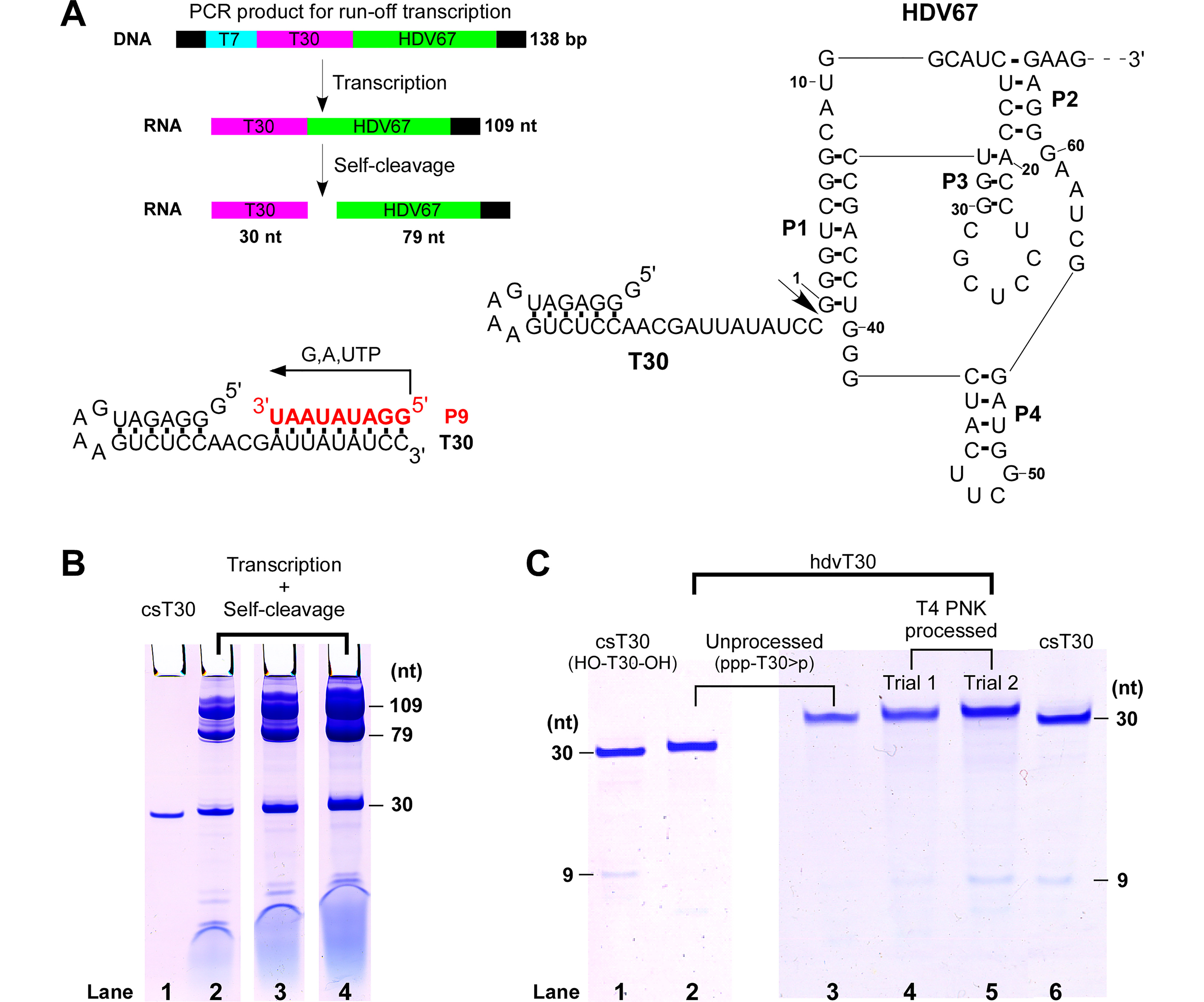
Figure 1. Preparation of a template RNA suitable for the WNV RdRP de novo synthesis through a HDV ribozyme-derived approach. (A) Top left: a diagram showing T7 RNAP transcription using the PCR product as the DNA template and transcript self-cleavage by the HDV ribozyme. The regions in cyan, magenta, and green indicate the promoter for T7 RNAP, T30, and HDV67, respectively. The black regions are the non-essential sequences introduced by the PCR primers. Bottom left: the sequences of the T30 template and the expected P9 product in the de novo RNA synthesis by the WNV RdRP in the presence of GTP, ATP, and UTP. Right: the secondary structure of the transcript including the T30 in the 5′-region and the HDV67 ribozyme in the 3′-region. The 3′ non-essential sequence is shown as a dashed line. The arrow indicates the cleavage site and HDV ribozyme nucleotide numbering starts from the cleavage site. The four base-paired regions of the HDV67 are labeled as P1–P4. (B) Denaturing polyacrylamide gel analysis of the T7 RNAP transcript and its self-cleavage products. Lane 1-csT30; lanes 2/3/4–1.5/3/6 μL transcription mixture. The 109-, 79-, and 30-nt species are the uncleaved transcript, HDV67-containing cleavage product, and T30, respectively. (C) Denaturing polyacrylamide gel analysis of de novo synthesis by the WNV NS5 RdRP. Templates used: lanes 1 and 6-csT30; lanes 2 and 3-hdvT30; lane 4, T4 PNK treated hdvT30 (trial 1); lane 5, T4 PNK treated hdvT30 (trial 2).
To generate the DNA templates for in vitro T7 RNAP transcription, PCR was carried out using the pRAV23-T30-HDV67 plasmid as the template and the 5′-GEN and 3′-HDV as the primers. The 138-base pair (bp) PCR product contains a T7 RNAP promoter immediately followed by the T30 and then by the HDV67 sequences (Figure 1A). In vitro run-off T7 RNAP transcription was carried out at 37 °C for 3.5 h in a solution containing 60 mmol/L HEPES (pH 7.8 by KOH), 2 mmol/L spermidine, 0.01% (vol./vol.) Triton X-100, 15 mmol/L dithiothreitol (DTT), 22 mmol/L MgCl 2, 4 mmol/L each NTP and 12.5 μg/mL T7 RNAP. The 109-nt transcripts were self-cleaved by HDV67 ribozyme, producing the 30-nt T30 RNA and the 79-nt HDV67-containing fragments (Figure 1A, 1B) at the end of the transcription reaction. The uncleaved transcript and both cleavage products were separated by 7 mol/L urea/20% polyacrylamide gel electrophoresis and visualized by Stains-All (Sigma-Aldrich) staining (Figure 1B). Homogeneous T30 RNA (hereafter named hdvT30) was obtained using this approach and its migration was slightly slower than the chemically synthesized T30 (csT30). Note that csT30 bears hydroxyl (-OH) groups at both the 5′- and 3′-ends (symbolized as HO-T30-OH), while the hdvT30 has a triphosphate at the 5′-end and a 2′, 3′-cyclic phosphate at the 3′-end (symbolized as ppp-T30 > p). The molecular weight and net charge differences between the two T30 constructs both contribute to their migration difference in the denaturing gel electrophoresis ( Figure 1B, 1C). The HDV cleavage efficiency is approximately 50% as judged by the band intensities of the uncleaved RNA and the cleavage fragments (Figure 1B, lanes 2-4), and the efficiency may be improved by multiple rounds of thermal cycling (Walker et al., 2003). Following a preparatory transcription and denaturing gel electrophoresis trial, the hdvT30 RNA was excised from the gel, electro-eluted by an EluTrap device (GE Healthcare), ethanol precipitated, self-annealed by snap cooling (3 min incubation at 95 °C and then cooled by ice), and stored at –80 °C in an RNA annealing buffer (RAB) containing 5 mmol/L Tris (pH 7.5 by HCl), 50 mmol/L NaCl, and 5 mmol/L MgCl 2.
WNV belongs to the Flavivirus genus and the Flaviviridae family. RdRPs from the Flaviviridae family initiate viral RNA genome replication in a primer-independent (also known as “de novo”) fashion (Kao et al., 2001). To prepare the WNV NS5 RdRP, the expression plasmid pET26b-WNV-NS5 (NS5 coding sequence consistent with GenBank asscession no. AF404756) was first transformed into E.coli BL21 (DE3) competent cells. Cells were grown at 37 °C in LB medium for about 10 h in the presence of 50 μg/mL kanamycin (KAN50) until OD 600 was 1.0, and were used to inoculate in 1 L LB medium with KAN50 for growth at 37 °C until OD 600 was 0.6. The cell culture was cooled to room temperature and protein expression was induced by 0.5 mmol/L isopropyl-β-D-thiogalactopyranoside (IPTG) at 25 °C for 12 h before harvesting. Cell lysis and protein purification were performed as described previously ( Lu and Gong, 2013), except that the gel filtration buffer was 5 mmol/L Tris (pH 7.5 with HCl), 300 mmol/L NaCl, 0.02% (wt./vol.) NaN 3, and 20% (vol./vol.) glycerol. The pooled fractions were supplemented with 5 mmol/L tris-(2-carboxyethyl)phosphine (TCEP), concentrated to about 15 mg/mL, flash frozen in liquid nitrogen as 10 or 20 μL aliquots, and stored at –80 °C. WNV NS5 protein concentration was determined by a calculated extinction coefficient of 221, 730 M –1cm–1 at 280 nm using the ExPASy ProtParam program (http://web.expasy.org/protparam/).
The de novo RNA synthesis by WNV NS5 was carried out at 35 °C for 60 min in an optimized condition of 50 mmol/L Tris (pH 7.5 with HCl), 25 mmol/L NaCl, 5 mmol/L MnCl 2, 4 mmol/L DTT, 300 μmol/L each GTP, ATP, and UTP, 4 μmol/L T30 RNA template and 6 μmol/L NS5. According to the sequence of T30, NS5 is able to initiate de novo RNA synthesis starting from the 3′-end of the template and produce a 9-mer RNA product (P9, ppp-GGAUAUAAU-OH) (Figure 1A). The csT30 and hdvT30 were used in parallel experiments. The P9 product was obtained using the csT30 construct but could hardly be detected when hdvT30 serving as the template. Because hdvT30 bore a 2′, 3′-cyclic phosphate at its 3′-end, we suspected that the cyclic phosphate group might inhibit NS5 de novo initiation by steric or electrostatic interferences in the finely controlled initiation catalysis (Appleby et al., 2015). To test this hypothesis, the bacteriophage T4 polynucleotide kinase (T4 PNK, New England BioLabs) was used to remove the cyclic phosphate of hdvT30 in a 3 h reaction carried out at 37 °C in a solution containing 40 mmol/L MES pH 6.0, 10 mmol/L MgCl 2, 5 mmol/L DTT according to the previously described methods (Das and Shuman, 2013). Two trials were performed with different amount of T4 PNK supplied per 1 nmol hdvT30 (trial 1, 10 units; trial 2, 20 units). After the dephosphorylation, the reaction mixtures were subjected to phenol/chloroform extraction and ethanol precipitation. The processed hdvT30 samples were dissolved in RAB and used as the RNA template in the de novo RdRP assay of the WNV NS5. The results demonstrated that the P9 product could be readily detected using the trial 1-derived T30 (Figure 1C, lane 4), while the amount of P9 obtained from trial 2-derived T30 was comparable to that obtained using the csT30 (Figure 1C, compare lane 5 and lane 6). These data suggest that the cyclic phosphate removal in trial 2 was quite efficient and the removal process is necessary in preparing the WNV NS5 de novo synthesis RNA template using the HDV-derived approach.
Taken together, we have successfully utilized the HDV ribozyme in the preparation of homogeneous RNA sample that may be free of sequence limitation at the 3′-end. To make the HDV approach-derived RNA suitable for directing de novo RNA synthesis by the WNV NS5 RdRP, the 2′, 3′-cyclic phosphate at its 3′-end needs to be removed. The cyclic phosphate interference with the WNV RdRP initiation is an interesting observation and the underlying mechanisms may deserve further investigation. The practical approach combining T7 RNAP transcription, HDV ribozyme self-cleavage, and T4 PNK 2′, 3′-cyclic phosphate removal utilized in this study is likely applicable for in vitro characterization of de novo viral RdRPs. In a broader context, this approach may be applicable for developing RNA-based assays and for X-ray crystallography-based or cryo-electron microscopy (EM)-based structural studies of RNA and RNA-protein complexes.
HTML
-
This work was supported by the National Key Basic Research Program of China (2013CB911100 to P.G.), the National Natural Science Foundation of China (31370198 and 31670154), the Open Research Fund Program of Wuhan National Bio-Safety Level 4 Laboratory of Chinese Academy of Sciences (NBL2017009), and the “One-Three-Five” Strategic Programs, Wuhan Institute of Virology, Chinese Academy of Sciences (Y605191SA1). We thank Dr. Bo Zhang for supplying the WNV NS5 expressing plasmid, Dr. Bo Shu, Dr. Guoliang Lu, and Peng Bi for helpful discussions, Liu Deng for laboratory assistance, and The Core Facility and Technical Support, Wuhan Institute of Virology, for access to instruments. The authors declared that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by the author.

 DownLoad:
DownLoad: