














HTML
-
The human immunodeficiency virus (HIV)/Acquired Immune Deficiency Syndrome (AIDS) epidemic, one of the most severe challenges in the field of public health, is still rampant worldwide. Approximately 38 million individuals live with HIV/AIDS nowadays (UNAIDS). Antiretroviral therapy (ART) can effectively inhibit HIV replication, and it has turned AIDS from a fast-fatal incurable disease to a manageable chronic disease. However, ART cannot fully cure HIV-related clinical symptoms, such as persistent inflammation, extensive immune activation, and abnormal lipid metabolism (Churchill et al. 2016; Koethe et al. 2020; Premeaux et al. 2020). Approximately 40% of HIV-infected patients suffer from dyslipidemia (Grand et al. 2020), resulting in an increased incidence of cardiovascular disease with a 1.5 to 1.7 times higher risk than that of the general population (Freiberg et al. 2013). Considering that an estimated 25.4 million AIDS patients are currently receiving life-long ART, it is becoming increasingly important to regulate lipid metabolism disorders in long-term ART-treated HIV patients. The American Infectious Diseases Association has added relevant items to the Guide for Primary Care of HIV-Infected Persons (Aberg et al. 2014). As a result, researchers are exploring novel antiviral drugs that can simultaneously reshape immune function, repress viral replication, and restore normal physiological metabolic balance, to achieve the coordinated control and the eventual elimination of HIV infection.
Cholesterol-25-hydroxylase (CH25H), also known as cholesterol-25-monooxygenase, belongs to the redox enzyme family, and it can catalyze cholesterol to produce 25-hydroxycholesterol (25HC), an oxidative product of cholesterol metabolism that is closely related to the regulation of lipid metabolism (Lund et al. 1998). CH25H and 25HC have been regarded as important regulators that maintain cholesterol homeostasis by inhibiting sterol regulator-binding protein (SREBP) and liver X receptor (LXR) (Lehmann et al. 1997; Radhakrishnan et al. 2004). Recently, CH25H and 25HC have also been shown to play critical roles in the regulation of inflammation, innate immunity, and subsequent adaptive immune responses through interferon signaling (Glass and Saijo 2010; Ludigs et al. 2012; Spann and Glass 2013; Zhao et al. 2020). Furthermore, CH25H and 25HC have been found to inhibit a variety of viruses, including Zika virus (ZIKV) (Li et al. 2017), HIV-1, human herpesvirus 1 (HSV-1), hepatitis C virus (HCV), Ebola virus (EBoV) (Liu et al. 2013; Doms et al. 2018), Lassa virus (Shrivastava-Ranjan et al. 2016), rabies virus (Yuan et al. 2019), and SARS-CoV-2 (Zu et al. 2020). Our recent study also indicated that 25HC treatment significantly suppressed simian immunodeficiency virus (SIV) infection by modulating both innate and adaptive immune responses in a dose-dependent manner in vitro, and enhanced SIV-specific IFN-γ-producing cellular responses but selectively suppressed proinflammatory CD4+ T lymphocytes in immunized mice with the SIV vaccine (Wu et al. 2018). Given the multifaceted functions of 25HC, this compound warrants exploration as a potential therapeutic drug in preclinical and clinical studies. Further, its antiviral immunity and therapeutic efficacy in a nonhuman primate model are unknown.
In the present study, we investigated how 25HC affects viral replication, lipid metabolism, and immune modulation in a chronically SIV-infected, long-term ART-treated RM model. We also explored the in vivo safety of 25HC in this nonhuman primate model. This study helps us understand the relationship between cholesterol metabolism, immune modulation, and antiviral activities by 25HC, and provides insight for developing novel therapeutic drug candidates against HIV infection and other related diseases.
-
25HC was kindly provided by Prof. Genhong Cheng and Feng Ma, and dissolved in ethanol, and stored at –20 ℃.
Reverse Transcriptase Inhibitors: (R)-9-(2-phosph-onylmethoxypropyl) adenine (PMPA, also called tenofovir) and beta- 2, 3-dideoxy-3-thia-5- uorocytidine (FTC, also called emtricitabine) were provided by Shanghai Desano Pharmaceutical Co., Ltd. These drugs were provided in powder formation of active pharmaceutical ingredient, and dissolved in 0.9% saline solution to a final concentration as below.
Peptide pools, which covered the entire SIVmac239 sequences of Gag, Pol, Env, Nef, Vif, Vpx, Vpr, Rev, and Tat proteins, were obtained through the AIDS Research and Reference Reagent Program, National Institutes of Health (NIH), USA. Peptide pools consisted of 15 amino acids shifted by 11 overlapping amino acids residues, and dissolved in dimethyl sulfoxide to a final concentration of 0.4 mg per peptide mL−1 before use. Concanavalin A (ConA, Sigma), ionomycin (Ion, Sigma) and phorbol myristate acetate (PMA, Enzo Biochem, Inc.) were prepared and stored according to the manufacturer's instructions.
The original SIVmac239 stock was obtained through the AIDS Research and Reference Reagent Program, National Institutes of Health (NIH), USA. The challenge stock was propagated and purified in peripheral blood mononuclear cells (PBMCs) of rhesus macaques (RMs) and tittered in 174×CEM cells in our lab.
-
Chinese RMs were housed in the Experimental Animal Center of Guangzhou Institutes of Biomedicine and Health (GIBH, Guangzhou, China). All procedures were performed by trained personnel under the supervision of veterinarians. Eleven RMs were recruited in this study and assigned to one of two treatment groups: 25HC combined with ART or ART-only. Briefly, all 11 RMs were intravenously inoculated with 5000 50% tissue culture infectious dose (TCID50) SIV at 0 days post infection (dpi) and received antiretroviral therapy (FTC/20 mg/kg/once daily + PMPA/30 mg/kg/once daily) from 69 to 132 dpi. During this period, 6 RMs in the 25HC+ART group received daily intravenous infusion of 25HC (1.5 mg/kg) from 119 dpi ~ 125 dpi, and the RMs in the ART-only group received intravenous drip vehicle (ethanol). To further study the therapeutic effect of 25HC alone, we also performed an additional week with 25HC treatment alone in these RMs of the 25HC + ART group, and the RMs of the ART-only group were given ethanol. Samples were collected to monitor the virological and immunological parameters following the experimental schedule, as shown in Fig. 1.

Figure 1. The experimental schedule and specimen collection in this study. A Eleven rhesus macaques were randomly assigned into two groups: the 25HC combined with ART group and the ART-only group. All rhesus macaques were intravenously injected with 5000 TCID50 (50% tissue culture infectious dose) SIV at day 0 (black arrow), and daily ART administration (blue shaded area) was started at 69 dpi (day post infection). The 6 macaques in the 25HC+ART group were intravenously dripped with 25HC (red shaded area) from 119 to 125 dpi, and the 5 rhesus macaques in the ART-only group were injected with vehicle (ethanol) at the same time. ART was interrupted at 132 dpi. At 154 dpi, the 25HC+ART group was infused with 25HC alone, and the ART-only group was injected with vehicle (ethanol). B Information on rhesus macaques enrolled in this study. C The schedule of specimen collection, including the time-points for SIV challenge, ART, 25HC treatment, plasma viral load detection, PBMC collection, blood biochemistry, complete blood count, and 25HC pharmacokinetic analysis. Dpi = days post infection.
-
PBMCs from SIV-infected RMs were isolated by standard Ficoll-Hypaque density gradient centrifugation, and then used for IFN-γ ELISPOT assay and intracellular cytokine staining (ICS) assay as we described previously (Sun et al. 2010; Pan et al. 2018). For IFN-γ ELISPOT assay, freshly isolated PBMCs were added at 4 × 105 cells/well in anti-RM IFN-γ monoclonal antibody (BD Pharmingen) pre-coated 96-well plates containing Immobilon-P membrane (Millipore, USA). SIV peptide pools were added into cells for 20–24 h for stimulation, and then a polyclonal anti-RM IFN-γ biotinylated detector antibody (BD Pharmingen) was added. The next day, the plates were washed and color was developed by incubating in NBT/BCIP (Pierce, Rockford, IL) for 10 min. Spots were counted under an ELISPOT reader (Bioreader 4000, BIOSYS, Germany), and data were reported as the number of spot-forming cells (SFCs) per million PBMCs. Concanavalin A stimulation was used as a positive control in the ELISPOT assay.
For ICS assay, 106 cells were stimulated with SIV peptides for 2 h, and then brefeldin A (BD Biosciences) was added for an additional 16 h. The cells were then washed and stained for 30 min with anti-CD3–Pacific Blue, anti-CD4–PE-CF594, anti-CD8–allophycocyanin (APC)–Cy7, anti-CD28–fluorescein isothiocyanate (FITC), and anti-CD95–phycoerythrin (PE)–Cy5. Next, the cells were suspended in 250 µL of Cytofix/Cytoperm solution (BD Pharmingen) for 20 min, washed with Perm/Wash solution (BD Pharmingen), and intracellularly stained with anti-IFN-γ–PE, anti-TNF-α–PE–Cy7 and anti-IL-2-APC (BD Pharmingen) for 30 min. Samples were analyzed with BD LSRFortessa™ (BD Biosciences) instrument and FlowJo software (Tree Star, Inc). PMA/I (phorbol myristate acetate + ionomycin) stimulation was used as a positive control in the ICS assay. The antibodies used in this study for analytical flow cytometry are listed in Supplementary Table S1.
For cell activation detection, PBMCs were detected with the following monoclonal antibodies: anti-CD3-PerCP, anti-CD4-FITC, anti-CD4-APC, anti-CD38-FITC, anti-CD25-APC, anti-CD69-PE, anti-CD95-PE-Cy5, anti-Ki67-PE, and anti-CCR5-PE. Samples were analyzed with an Accuri C6 flow cytometer (BD Biosciences) and FlowJo software (version 10; Tree Star, Inc.).
-
The SIV RNA copies in plasma were quantitated by real-time PCR as described previously (Wong et al. 1997; Kimata et al. 2016). Briefly, viral RNA was extracted from plasma using the QIAamp Viral RNA Minikit (Qiagen), and then quantitated using the QuantiTect SYBR Green RT-PCR Kit (Qiagen) with LightCycler480 II Real-time PCR system (Roche). Primers were designed to match the SIVmac239 gag sequence. The copy number of viral RNA was calculated based on the standard curve of an in vitro-transcribed fragment of the SIVmac239 gag gene. The limitation for this assay was 100 copies mL−1 plasma. Sequences of all primers used in this study were listed in Supplementary Table S2.
-
Peripheral blood and plasma samples were collected following standard protocols, and the complete blood count and biochemical tests were performed with Sysmex automatic modular animal blood and body fluid analyzer XN-1000 V and Hitachi Automatic Aralyzer 3100 by Guangdong Laboratory Animals Monitoring Institute.
To study the preliminary pharmacokinetics of 25-hydroxycholesterol, lipids were extracted from plasma samples using an improved Bligh/Dyer extraction method (two extractions). The lipids were dried in a vacuum rotary thickener and then remelted in an ethanol and isotope mixture containing 1% (W/V) butylated hydroxytoluene (BHT). The lipids were incubated at 37 ℃ for 1 h, deionized water and ethanol were added, and the samples were incubated for another 15 min. Subsequently, the upper organic phase was obtained by centrifugation, and pyridine acid derivatization was carried out. After derivatization, a liquid-mass spectrometer (Exion UPLC-QTRAP 6500 PLUS, Sciex) was used for analysis, and electrospray ionization (ESI) mode was used for quantitative analysis. The isotopic internal standard is d6-25-hydroxycholesterol.
-
Flow cytometry software analysis was performed using FlowJo 10 (Tree Star Inc.). Graphical representations were generated with GraphPad Prism 8 (GraphPad Software Inc., La Jolla, CA) and IBM SPSS Statistics 25.0. Two-tailed P values were calculated, and differences were considered as statistically significant when P < 0.05.
Drugs, SIV Peptide Pools, and Virus
Animals and Ethical Statement
Immunological Assays
Viral Load Determination by Real-Time PCR
The Complete Blood Count, Blood Biochemical Test, and the Preliminary Pharmacokinetics of 25HC in Rhesus Macaques
Data Analysis
-
To explore the effect of 25HC on the SIV-infected RM model, we designed the following experiments (Fig. 1A). Eleven RMs were recruited and assigned into two groups based on body weight (4–13 kg), sex, age (6–15 years old), and initial immune status (Fig. 1B): 25HC combined with ART administration group (25HC + ART), and ART-alone administration group (ART-only). All 11 RMs were intravenously inoculated with 5000 50% tissue culture infectious dose (TCID50) SIV at 0 days post infection (dpi), and received antiretroviral therapy (FTC/20 mg/kg/once daily + PMPA/30 mg/kg/once daily) from 69 to 132 dpi. During this period, 6 RMs in the 25HC + ART group received daily intravenous infusion of 25HC (1.5 mg/kg) from 119 dpi ~ 125 dpi, and the RMs in the ART-only group received intravenous infusion of vehicle (ethanol). To study the therapeutic effect of 25HC alone, we performed an additional week of 25HC administration. The experimental operation and specimen collection of RMs in this study are shown in Fig. 1C.
-
Enzymatic product 25HC has been reported to inhibit a variety of viruses (Zhao et al. 2020). Thus, we sought to evaluate its effect on SIV replication in the RM model. At 56 dpi, the median level of plasma viral load in all RMs was 4.3 lg (copies/mL) (range 3.9–5.4 lg). After initializing ART at 69 dpi, the plasma viral load in all RMs rapidly decreased until it was undetectable (Fig. 2A, 2B). Interestingly, at 126 dpi (the 58th day of ART), the plasma viral load of two RMs in the ART-only group rebounded to 4.7 and 3.9 lg (copies/mL) respectively, accounting for 40% of animals in this group. However, only one RM in the 25HC+ART group rebounded to 4.0 lg (copies/mL), accounting for 16.67% of animals in this group (Fig. 2C). There was no statistically significant difference in the viral rebound ratio between the two groups, possibly due to the small number in this study.
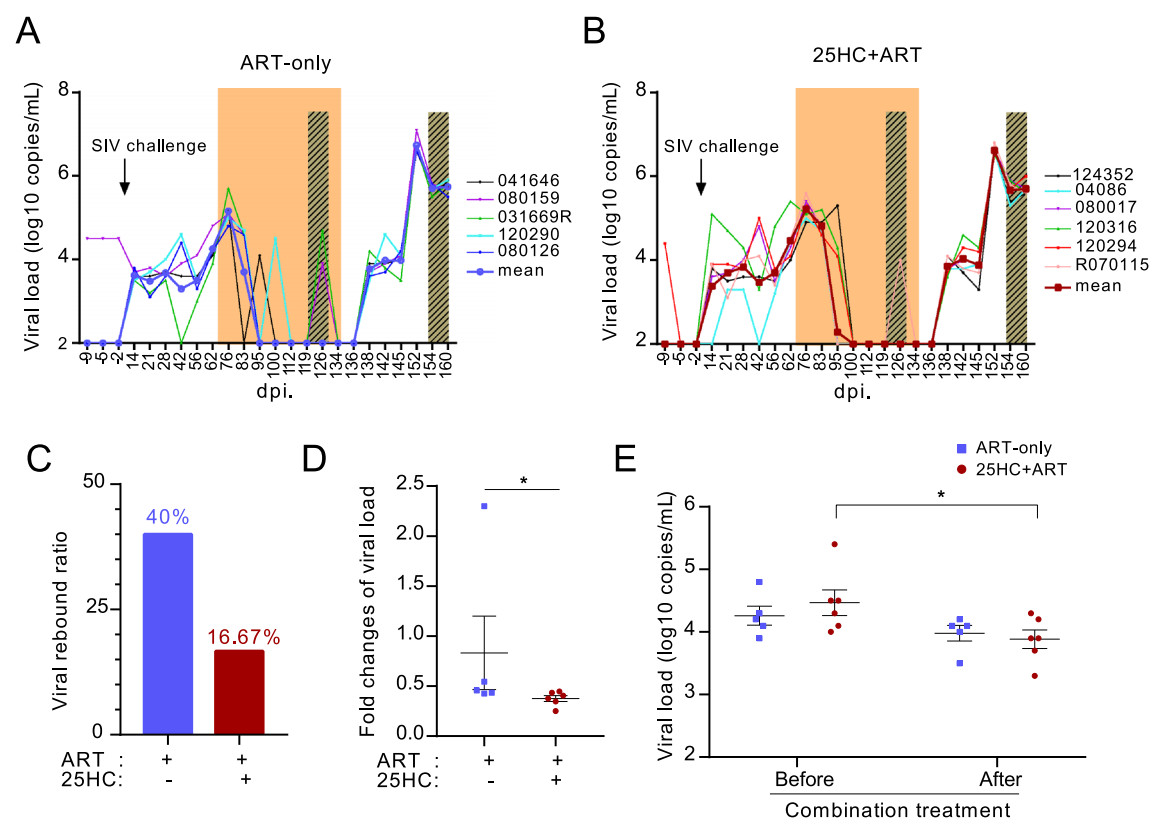
Figure 2. 25HC combined with ART improved the control of viral replication in SIV-infected RMs. A, B Plasma viral load in the ART-only group (n = 5) and 25HC + ART group (n = 6) was measured by one-step quantitative RT-PCR during the whole period. A log10 transformed value of RNA copies per mL is shown. C The ratio of viral rebound during ART between the two groups. D The fold changes of peak plasma viral load (VL) relative to baseline (VL83dpi) between the two groups. P values were obtained from Mann–Whitney test. E The variation in plasma viral load between the two groups before (62 dpi.) and after combination treatment (145 dpi.). P values were obtained from paired t test. The area shaded in orange represents ART administration, and the area shaded in cross grains represents the treatment of 25HC alone or vehicle. All average data are presented as the mean ± SEM. *, P < 0.05.
The plasma viral load of all RMs was monitored before ART and after ART termination. We noticed that the fold changes of peak plasma viral load relative to baseline were lower in the 25HC+ART group (Fig. 2D). In addition, the average set-point of plasma viral load in the 25HC + ART group was 4.5 lg (copies/mL) before treatment, and rebounded to 3.9 lg (copies/mL) after drug withdrawal, which was significantly lower than the viral load before treatment. However, there was no significant change between plasma viral load before ART and after ART termination in the ART-only group (Fig. 2E). We next wondered whether 25HC treatment alone could also play a role in controlling plasma viral load (Fig. 1A), but we found no significant change in plasma viral load with or without 25HC treatment alone (Fig. 2A and 2B). Overall, these data indicated that 25HC alone may not be sufficient to suppress the viremia in SIV-infected RMs, but 25HC combined with ART may be helpful in controlling viremia at a lower level after viral rebound in RMs.
-
We then determined how 25HC affects the quantity and quality of T cells in vivo in the RM model. The ratio of CD4/CD8 is considered to be an important indicator in predicting the progression of HIV disease (Spann and Glass 2013). The high baseline CD4/CD8 ratio was associated with immune reconstitution success (Mussini et al. 2015). Interestingly, we found that the CD4/CD8 ratio of RMs in the 25HC + ART group was significantly increased, and higher than that of the ART-only group (Fig. 3A) (Supplementary Table S3). These results showed that 25HC combined with ART restores the CD4/CD8 ratio, which is helpful for immune reconstitution.

Figure 3. 25HC combined with ART restored the ratio of CD4/CD8 lymphocytes and regulated the differentiation of T lymphocytes in SIV-infected RMs. To assess the T cell number in these SIV-infected RMs, the true T cell count was measured by flow cytometry. A The ratio of CD4/CD8 in SIV-infected rhesus macaques before (95 dpi) and after (161 dpi) getting combination treatment. P values were obtained from Tukey correction for multiple comparisons analysis. B, C, D Expression levels of CCR5, CD69, and Ki67 in CD4+ T cells before (95 dpi) and after (152 dpi) RMs were received combination treatment. P values were obtained from paired t test analysis. E Dynamic proportion change of naïve, Tcm and Tem CD4+ T cells in PBMCs from RMs against SIV gag stimulation in this study. P values were obtained from unpaired t test. F Dynamic proportion change of naïve, Tcm and Tem to CD8+ T cells against SIV gag stimulation along this study. P values were obtained from unpaired t test. Tcm represents central memory T cell, and Tem represents effective memory T cell. All average data are presented as the mean ± SEM. (*P < 0.05, ** P < 0.01).
It is generally considered that the hyperactivation of CD4+ T cells might provide more susceptible targets for HIV-1 acquisition. To investigate the activating status of CD4+ T cells by 25HC, we detected the expression of CCR5, CD69, and Ki67 on the surface of CD4 cells. In this study, there was a trend towards reduced expression of CCR5 on the surface of CD4+ T cells in RMs that received both 25HC combined with ART and ART-only treatment. Notably, the average frequency of CCR5/CD4 in RMs of the 25HC + ART group decreased from 7.9% to 4.8% after getting combined treatment of 25HC and ART (Fig. 3B). The expression of CD69 on the surface of CD4+ T cells in RMs was significantly decreased after 25HC combined with ART (Fig. 3C), suggesting that 25HC intervention may help reverse the overactivation of CD4+ T cells. Moreover, the expression of Ki67 on the CD4+ T cell surface significantly decreased in RMs that received 25HC combined with ART and ART-only (Fig. 3D). These findings suggest that 25HC combined with ART may reverse the overactivation of CD4+ T cells in SIV-infected RMs.
We also determined whether the 25HC intervention can affect the activation state of CD8+ T cells. The expression of CD38, PD-1, and HLA-DR on the surface of CD8+ T cells was detected, and we did not find a statistically significant difference between the two groups of RMs (Supplementary Fig. S1E).
We then detected the frequency of naïve T cell subsets, central T memory subsets (Tcm), and effector T memory subsets (Tem), based on the expression levels of CD28 and CD95 as described in our previous study (Sun et al. 2013; Wu et al. 2018). In this study, there was a similar dynamic of the proportion of these three subsets in CD4+ and CD8+ T cells in SIV-infected RMs receiving ART with or without 25HC (Fig. 3E, 3F) (Supplementary Fig. S1A–1D).
-
We next investigated whether 25HC therapy modulated the immune response in SIV-infected RMs, especially the antigen-specific cellular immune response. The frequency of SIV antigen-specific IFN-γ-secreting cells, determined by IFN-γ enzyme-linked immunospot assay (ELISPOT), was significantly increased in the RMs of the 25HC-combined-with-ART group compared to those of the ART-only group (Fig. 4A). Further analysis showed that these IFN-γ-mediated ELISPOT immune responses in the group of 25HC combined with ART were significantly enhanced against SIV Pol, Vpx, Vpr, Vif, Nef, Rev, and Tat peptide stimulation (Fig. 4B), while there was no significant change of immune responses against the above-mentioned peptides in the ART-only group (Fig. 4C). These data indicated that administration of 25HC during ART enhanced total IFN-γ expression and might contribute to its broad-spectrum antiviral function.

Figure 4. 25HC combined with ART improved SIV antigen-specific IFN-γ production, and selectively attenuated proinflammatory cytokine secretion in RMs. A The frequency of SIV antigen-specific IFN-γ-secreting cells was determined by IFN-γ ELISPOT assay before (119 dpi.) and after combination treatment (154 dpi.). P values were obtained from paired t test analysis. B, C Antigen-specific immune responses against different SIV antigen stimulations in SIV-infected rhesus macaques. P values were obtained from paired t test analysis. D, E The percentage of single-, double-, and triple-cytokine secretion by CD4+ and CD8+ T cells under SIV gag/pol/env stimulation before (42 dpi.) and after combination treatment (138 dpi) in two groups. F The percentage of TNF-α secreted by CD4+ T cells under SIV pol stimulation before (42 dpi) and after combination treatment (138 dpi) in 25HC+ART group. P values were obtained from Tukey's multiple comparisons test. G The proportion of IL-2 secreted by CD8+ T cells under SIV pol and env stimulation before (42 dpi.) and after combination treatment (138 dpi) in 25HC + ART group. P values were obtained from Tukey's multiple comparisons test. All average data are presented as the mean ± SEM. (*P < 0.05, ** P < 0.01, *** P < 0.001)
To assess how 25HC affects antigen-specific CD4+ T and CD8+ T cells, peripheral blood mononuclear cells (PBMCs) were isolated from RMs, and the polyfunctionality of T lymphocytes was assessed using multiparameter intracellular cytokine staining (ICS). The proportion of double and triple cytokine-secreting CD4+ and CD8+ T cells was increased in all the RMs after therapy (Fig. 4D, 4E). Interestingly, the frequency of TNF-α-secreting CD4+ T cells and IL-2-secreting CD8+ T cells was significantly decreased in RMs treated with 25HC combined ART (Fig. 4F, 4G), suggesting that 25HC may have an effect on suppressing proinflammatory cytokines, such as TNF-α and IL-2 (Upadhyay et al. 2020). These data demonstrated that 25HC combined with ART could enhance the total cellular responses but inhibit the proinflammatory responses by selectively suppressing the secretion of IL-2 and TNF-α in SIV-infected RMs.
-
While 25HC has been reported to play a critical role in the regulation of cholesterol (CHO) metabolism in vitro, there is a lack of related information in vivo. Thus, we examined how 25HC administration affected cholesterol-related lipid changes in SIV-infected RMs. After 25HC treatment, the concentration of high-density lipoprotein cholesterol (HDL-c) in RM plasma increased (Fig. 5A, 5B), and concentrations of CHO and low-density lipoprotein cholesterol (LDL-c) were significantly decreased (Fig. 5C, 5D). These results indicated that 25HC treatment did not increase the risk of atherosclerosis in SIV-infected RMs. In contrast, it may help to reverse the disorder of lipid metabolism caused by HIV/SIV infection.

Figure 5. The in vivo safety and preliminary pharmacokinetics of 25HC in SIV-infected RMs. A, B, C, D The dynamic changes in triglyceride (TG), high-density lipoprotein cholesterol (HDL-c), cholesterol (CHO) and low-density lipoprotein cholesterol (LDL-c) concentrations in SIV-infected RMs during this study. P values were obtained from unpaired t test. E, F, G The concentrations of alanine aminotransferase (ALT), aspartate aminotransferase (AST), and the ratio of ALT/AST in the plasma of RMs during this study. H, I The concentrations of creatinine (CREA) and blood urea nitrogen (BUN) in the plasma of RMs during this study. J, K The concentrations of lactate dehydrogenase (LDH) and glucose (GLU) in the plasma of RMs during this study. The contents of white blood cells (WBCs) (L), red blood cells (RBCs) (M), hemoglobin (HGB) (N), and platelets (PLTs) (O) in SIV-infected RMs during this study. P values were obtained from unpaired t test analysis (E–O). P The concentration of 25HC in the plasma of RMs at various time points after 25HC intervention. The P value was obtained from the Kruskal–Wallis test and Dunn's test. The orange-shaded area represents the ART administration, and the area shaded in the cross grain represents the treatment with 25HC alone or vehicle. All average data are presented as the mean ± SEM. (*P < 0.05).
We then evaluated the potential toxicity of 25HC to RMs throughout the study, including hepatic (Fig. 5E–5G), renal (Fig. 5H, 5I), and cardiotoxicity (Fig. 5J) effects. Throughout the course of the study, the change trend of the potential renal and cardiotoxicity index of RMs in the 25HC+ART group was similar to that in the ART-only group. There was no significant difference between the two groups (Fig. 5E–5K), indicating that 25HC will not cause additional toxicity to the kidney and heart. Compared to the ART-only group, the ratio of AST/ALT in RMs of 25HC+ART group was within the normal range (Yu et al. 2019), although there was a higher concentration of aspartate aminotransferase (AST) in the 25HC+ART group (Fig. 5F), suggesting that 25HC will not cause additional toxicity to liver. The observed higher AST concentration in the 25HC+ART group might be due to RMs' individual differences, since the concentration of AST in the 25HC+ART group was higher at the beginning of this study (42 dpi). These results demonstrated that daily infusion of 25HC (1.5 mg/kg) for one week did not cause detectable toxicity in SIV-infected RMs.
We also conducted a complete blood cell count over the course of the entire study, and the levels of white blood cells (WBCs), red blood cells (RBCs), hemoglobin (HGB), and platelets (PLTs) in RMs that received 25HC combined with ART were all within the normal reference value range after treatment (Fig. 5L–5O). However, we found that after RMs treated with 25HC combined with ART (152 dpi), the level of lymphocyte (LYM), ratio of LYM to WBC (LYM/WBC), monocyte (MONO), ratio of MONO to WBC (MONO/WBC), and basophil (BASO) were significantly lower than that of RMs who received ART alone (Supplementary Fig. S2A–S2F). The level of neutrophils (NEU), ratio of NEU to WBC (NEU/WBC), eosinophil (EOS), and ratio of EOS to WBC (EOS/WBC) were significantly higher than that of RMs who received ART alone (Supplementary Fig. S2G–S2J). Although the lymphocyte value of the 25HC+ART group was lower than the normal standard, the rate of decrease in the lymphocyte value before (95 dpi) and after (126 dpi) combined treatment was lower than that of the ART-only group, indicating that the number of lymphocytes in the 25HC+ART group was more stable. The level of monocytes (MONO), and the ratio of MONO to WBCs (MONO/WBCs) were all in the normal range. The concentration of hematocrit (HCT), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), red cell distribution width (RDW), and mean platelet volume (MPV) in peripheral blood of all RMs were almost no different between the two groups (Supplementary Fig. S2K–S2P), but MCV and MCHC deviated from the normal value, suggesting that chronically SIV-infected RMs may have anemia (Lee et al. 2012). These data showed that 25HC had no significant effect on the cell composition of peripheral blood in SIV-infected macaques.
To track the preliminary pharmacokinetics of 25HC in RMs, we collected blood from 6 RMs at different time points after they received 25HC intravenous infusion and monitored the changes in 25HC concentration (Fig. 5P). We found that the concentration of 25HC in plasma peaked one hour after intravenous infusion, then gradually decreased and fell to almost the same level as the initial concentration 3 h after 25HC intervention. After 24 h of drug intervention, the concentration of 25HC in plasma was at a low level, and it was maintained for at least 72 h after treatment (Fig. 5P).
Animal Model and Experimental Design
25HC Combined with ART Might be Helpful in Controlling Viremia in SIV-Infected RMs
25HC Combined with ART Restored the Ratio of CD4/CD8 Lymphocytes and Regulated the Differentiation of T Lymphocytes in SIV-Infected RMs
25HC Combined with ART Improved SIV Antigen-Specific IFN-γ Production, and Selectively Attenuated Proinflammatory Cytokine Secretion in RMs
Assessment of the In Vivo Safety and Preliminary Pharmacokinetics of 25HC in SIV-Infected RMs
-
Recently, increasing data have supported the broadly antiviral activities of CH25H and 25HC (Radhakrishnan et al. 2004; Liu et al. 2013; Li et al. 2017; Song et al. 2017; Wu et al. 2018). However, the potential therapeutic effect of 25HC in SIV-infected RMs remains to be studied. Thus, we evaluated the effectiveness and safety of 25HC in a chronically SIV-infected RM model for the first time.
Anti-inflammatory therapy is helpful for immune reconstruction in chronically HIV-1 patients, due to persistent inflammation and abnormal immune activation during HIV-1 infection. However, the exact roles of 25HC in modulating the inflammatory responses are still under extensive investigation. Some studies reported that 25HC inhibited the production of interleukin-1 family cytokines and inflammatory body activity (Reboldi et al. 2014; Dang et al. 2017; Ouyang et al. 2018), but another study found that 25HC promoted the secretion of inflammatory cytokines (Gold et al. 2014). In this study, our in vivo data in SIV-infected RMs showed that the proinflammatory secretion of SIV-specific IL-2 and TNF-α was selectively inhibited after 25HC treatment. Interestingly, there was no significant effect on the secretion of SIV-specific IFN-γ by CD4+ and CD8+ T cells, which has a direct antiviral function (Bovolenta et al. 1999; Papasavvas et al. 2019). Given that no significant change was found in the secretion of SIV-specific IFN-γ by CD4+ T cells and CD8+ T cells after 25HC treatment, we hypothesized that the enhancement of SIV-specific IFN-γ by ELISPOT assay might be mainly released by other innate immune cells including natural killer (NK) cells and macrophages etc., and the IFN-γ secretion by these innate immune cells was thought to be associated with delayed disease progression (Jiang et al. 2013). Consistent with our hypothesis, it was reported that 25HC induced the IFN-γ expression by macrophages in a liver X receptor (LXR)-dependent manner, and the increased IFN-γ subsequently promoted CH25H expression (Liu et al. 2018) and 25HC production (Diczfalusy et al. 2009). In addition, CD4+ T cells are the main target of HIV/SIV infection (Clapham et al. 1993); therefore, chronic HIV-1 infection and disease progression are usually accompanied by a decrease in the number of CD4+ T cells and the ratio of CD4/CD8 (Maina et al. 2015; Bruno et al. 2017; Mutoh et al. 2018). Of note, we found that the ratio of CD4/CD8 was significantly increased in SIV-infected RMs after receiving 25HC combined with ART. Overall, these immunological benefits are represented as a good predictor to control the progression of AIDS.
Reminiscent of the ongoing coronavirus disease 2019 (COVID-19) pandemic worldwide, lymphopenia and inflammatory cytokine storms are often observed in highly pathogenic coronavirus infections(such as severe acute respiratory syndrome (SARS), Middle East respiratory syndrome coronavirus (MERS), and COVID-19), and are related to the severity of the disease (Chen et al. 2020; Wang et al. 2020). However, how different lymphocyte subpopulations and inflammatory cytokine dynamics change in peripheral blood during SARS-CoV-2 infection remains largely unclear (Cuadrado et al. 2020; Liu et al. 2020). The anti-inflammatory role of 25HC may have effects on the inhibition of inflammatory cytokines storm in COVID-19 patients. In fact, our recent study showed that 25HC was upregulated in severe COVID-19 patients, and 25HC treatment inhibited SARS-CoV-2 infection in vitro and in humanized ACE2 mice (Zu et al. 2020). As a result, further exploration of the therapeutic effect of 25HC in COVID-19 patients is warranted.
In part due to its ability to inhibit cholesterol biosynthesis (Brown et al. 1975; Brown and Goldstein 1997; Adams et al. 2004), 25HC is considered an important antiviral molecule (Schoggins and Randall 2013; Lv et al. 2019). Manipulating cellular cholesterol levels is an important process in host and virus interactions. HIV-1 patients receiving long-term antiretroviral therapy are usually observed to have dyslipidemia (Maggi et al. 2017), and a recent study reported that the lipid mass spectrum was significantly changed in HIV-1 patients (Koethe et al. 2020). Compared with healthy people, HIV-1 patients have higher levels of triglycerides and total cholesterol, while high-density lipoprotein (HDL) cholesterol levels are lower (Bernal et al. 2008), leading to an increased incidence of cardiovascular disease. Epidemiological data have confirmed an independent positive association between LDL-c and cardiovascular disease risk (Chen et al. 1991; Stamler et al. 1993), and the level of cholesterol is strongly associated with the risk of atherosclerosis and cardiovascular disease (McGill et al. 2008; Gidding et al. 2016; Ference et al. 2017). Our study showed that the concentration of plasma HDL-c was increased, and the concentrations of cholesterol and LDL-c were significantly decreased when RMs were receiving 25HC treatment. These results indicated that 25HC intervention may have an effect on inhibiting cholesterol biosynthesis, and thus might reduce the risk of cardiovascular disease in HIV-1 chronically infected patients. In addition, 25HC was metabolized within three hours in RMs, and its concentration decreased to a much lower level than that before treatment at 24 h, and this continued at least until 72 h after administration. Due to the complexity of cholesterol metabolism (Hewing and Landmesser 2015), the reasons for this observation need to be further examined. Our findings provide a new way to control HIV-1 infection by regulating cholesterol metabolism.
This study has some limitations. First, RMs recruited in this study had a wide range of ages. To minimize the possible effect of age, we divided these animals into two groups based on average age and the initial immune status to SIV antigens. Second, only 11 RMs were enrolled in this study, and the animal number may be enlarged in future studies to confirm our results. In this study, we found that 25HC combined with ART was helpful in controlling viremia at a lower level after viral rebound in SIV-infected RMs. Our data also showed that 25HC alone may not be sufficient to suppress viremia in SIV-infected RMs. The exact mechanism for this observation is still unknown. One possible explanation might be the low dosage of 25HC employed in this study. Because of the relatively small number of experimental animals, we did not study the dose-dependent effectiveness of 25HC in this study, but under the current conditions, 25HC treatment was effective when combined with ART, possibly because of a synergistic interaction. Importantly, our data also showed that the 25HC compound has potential druggability with in vivo safety and preliminary pharmacokinetics studies. Nevertheless, given the multifaceted functions of 25HC in inhibiting viral infection, promoting lipid metabolism, and modulating immunity, we should further examine the potential of this drug in preclinical and clinical studies.
In summary, our results suggested that the regimen of 25HC combined with ART provided a profound modulation of virological and immunological benefits in chronically SIV-infected, ART-treated RMs, including suppressing viral rebound, enhancing SIV-specific cellular immune responses, restoring the CD4/CD8 T cell ratio, and inhibiting proinflammatory cytokine secretion in a nonhuman primate model (Fig. 6). This study helps explain the antiviral activity of 25HC and provides new insights into the development of novel immunotherapeutic strategies against HIV-1 infection and other related diseases.
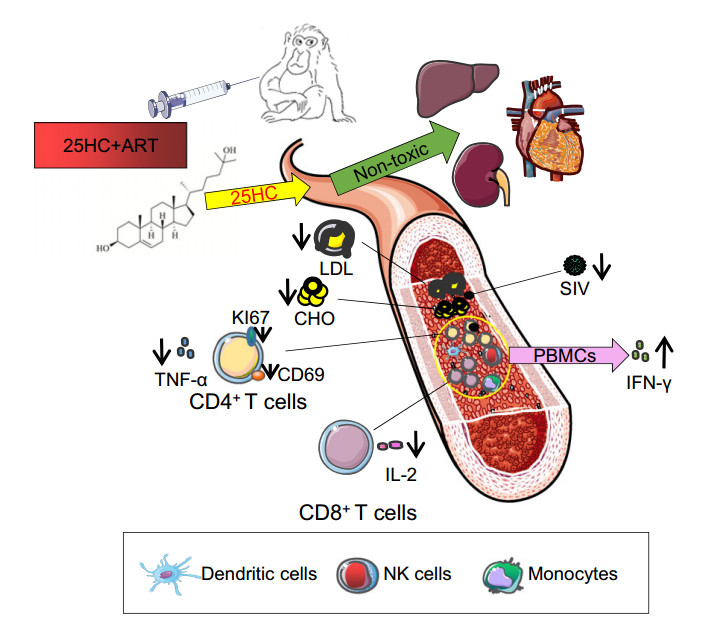
Figure 6. Pattern to illustrate the role of 25HC in controlling SIV infection. In this study, 25HC showed no toxicity to the liver, heart or kidney in SIV-infected RMs. The 25HC + ART regiment promoted the secretion of SIV-specific IFN-γ but suppressed the secretion of inflammatory cytokines, reversed the hyperactive state of CD4+ T cells, and restored CD4/CD8 T cell ratio in SIV-infected RMs. Furthermore, 25HC combined with ART reduced the concentration of total cholesterol and low-density lipoprotein in plasma. Overall, these immunological and metabolic benefits are helpful in controlling viral replication and suppressing the progression of AIDS.
-
We thank Yichu Liu and Xiangjie Feng for technical assistance in the RMs experiment. We appreciate the NIH AIDS Research and Reference Reagent Program for providing SIV peptide pools. This work was supported by the National Natural Science Foundation of China (81971927, 31870912, 32000124), the National Science and Technology Major Project of China (2018ZX10731101-002), the National Key Research and Development Program of China (2018YFA0900803), the Science and Technology Planning Project of Shenzhen City (20190804095916056, JCYJ20200109142601702), the High Level Project of Medicine in Longhua, Shenzhen (HLPM201907020105), China Postdoctoral Science Foundation (Grant No. 2019M663140), and the Municipal Health and Medical cooperation innovation Major Project of Guangzhou City (201704020219, 201803040002).
-
CS and PL designed and supervised the project; CW, JZ, RL, and CS collected literature and drafted the original manuscript; CW, JZ, RL, PL, RH, and YH performed the experiment; CW, JZ, and RL analyzed the data; HY, GC, FM, and LC contributed the materials and reagents. All authors have read and agreed to the final version of the manuscript.
-
This study was carried out in accordance with the ''Regulations for the Administration of Affairs Concerning Experimental Animals'' by the State Council of the People's Republic of China, and the protocol was approved by the Institutional Animal Care and Use Committee of GIBH (IACUC Permit Number: 2019052).
-
The authors declare that they have no conflicts of interest. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.

 DownLoad:
DownLoad: