












HTML
-
Zika virus (ZIKV) is a mosquito-borne RNA virus, which belongs to the family Flaviviridae (Simmonds et al. 2017). It was initially isolated from Rhesus monkey in Uganda in 1947 and was sporadically reported in humans since then (Dick et al. 1952). In 2007, the first epidemic of ZIKV occurred in Yap Island of Micronesia, and the symptoms of infected individuals are generally mild and often asymptomatic (Duffy et al. 2009). ZIKV subsequently spread to French Polynesia in 2013, where patients with GuillainBarre x syndrome were reported (Oehler et al. 2014). In 2015–2016, a large outbreak of ZIKV occurred in Central and South American countries with millions of people infected (Campos et al. 2015). Contrary to the mild symptoms in the past, the recently isolated ZIKV strains could cause severe neurological defects in neonates, such as microcephaly (Brasil et al. 2016). With widespread social panic, this epidemic was declared as a global health emergency by World Health Organization in 2016 (WHO 2016). Although the epidemic of unprecedented magnitude in the Americas has ended, the recent outbreak in India indicates that ZIKV will continue to be a public health threat in the near future (Sharma et al. 2020).
Numerous studies have tried to investigate the potential causes for ZIKV-related microcephaly, and viral infectioninduced cell cycle perturbation and cell apoptosis are believed to be the main reason for microcephaly (Ghouzzi et al. 2016; de Sousa et al. 2018). ZIKV preferentially targets human neural progenitor cells (hNPCs) during infection (Tang et al. 2016). And ZIKV infection rapidly induces cleavage of caspase-3, 8 and 9 and leads to apoptotic cell death of hNPCs, which can be prevented by pan-caspase inhibitors (Souza et al. 2016). Caspase-3- mediated apoptosis also occurs in ZIKV-infected 3D brain organoids and animal models (Li et al. 2016; Pettke et al.2020). Moreover, the brain tissues from the ZIKV-infected fatal cases with microcephaly showed positive staining of apoptosis markers, such as Bax, BCL2 and caspase 3 (de Sousa et al. 2018), further implying the link between neural cell apoptosis and ZIKV-induced microcephaly.
The tumor suppressor p53 protein plays an important role in cell apoptosis during brain development (Marjanovic et al. 2015; Little and Dwyer 2019). Normally, the p53 is a short-lived protein and keeps a low level in cells, which is regulated by the E3 ubiquitin ligase MDM2 (Kubbutat et al.1997). In response to a range of cellular stress, p53 protein is stabilized and activated by phosphorylation to stimulate the transcription of a series of target genes, including cyclindependent kinase inhibitor p21, pro-apoptotic protein Bax and the BH3-only proteins puma and Noxa (Beckerman and Prives 2010). These p53 target genes regulate multiple biological processes, including apoptosis, cell cycle arrest and senescence (Bieging et al. 2014). Abnormal activation of p53 signaling pathway has been reported to promote hNPC apoptosis and lead to microcephaly during fetal brain development (Toki et al. 2018), and deletion of endogenous p53 could rescue the microcephaly phenotype in animal models (Marjanovic et al. 2015; Breuss et al. 2016; Bianchi et al. 2017). Moreover, activation of p53 pathway in ZIKVinfected hNPCs has been recently documented (Ghouzzi et al. 2016) and p53-mediated apoptosis has been speculated as one of the main causes of ZIKV-induced microcephaly phenotype (Zhang et al. 2016). However, the detailed molecular mechanism to activate the p53-mediated apoptosis in ZIKV-infected hNPCs is still not fully understood.
As a single-stranded positive-sense RNA virus, the ZIKV genome contains one open reading frame and highly structured 50 and 30 untranslated regions. The encoded single polyprotein can be further cleaved into ten functional proteins, including three structural proteins (envelope, precursor membrane, and capsid) and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5). In this study, we found that, among the ten viral proteins, NS5 could most significantly activate the p53 signaling pathway. NS5 directly interacted with the CTD domain of p53 and then increased its stability in nuclei. Intriguingly, the expression of ZIKV-NS5 alone in hNPCs could induce the p53-mediated apoptosis, implying its contribution to the microcephaly caused by ZIKV infection.
-
ReNcell CX cells, a cell line of human neural progenitor cells (hNPCs), were cultured in ReNcell NSC maintenance medium (SCM005, Millipore) with 20 ng/mL FGF-2 (100- 18B-100, PeproTech) and 20 ng/mL EGF (315-09-100, PeproTech) in 20 lg/mL laminin-coated dishes at 37 ℃ with 5% CO2. HEK293T cells and Vero cells were cultured in Dulbecco's Modified Eagle Medium (DMEM) and supplemented with 10% fetal bovine serum (FBS) at 37 ℃ with 5% CO2. Cell lines were tested for mycoplasma.
-
Primary antibodies used in this study were listed: mouse monoclonal antibody anti-FLAG (F1804, Sigma-Aldrich), rabbit polyclonal antibodies including anti-GFP (G1544, Sigma-Aldrich), anti-Zika NS5 protein (GTX133329, GeneTex), anti-p21 (10355–1-AP, Proteintech), antiGAPDH (10494-1-AP, Proteintech), anti-puma (55120-1- AP, Proteintech), anti-p53 (10442-1-AP, Proteintech), antiGST (10000-0-AP, Proteintech), anti-HA (51064-2-AP, Proteintech), anti-PARP1 (13371-1-AP, Proteintech), and anti-phospho-p53(Ser15) (9284, Cell Signal Technology). Secondary antibodies: donkey anti-rabbit IgG (H + L) Alexa Fluor 555 (A31572) for immunoblotting and goat anti-mouse IgG (H + L) (31430) and Goat anti-rabit IgG (H + L) (31460) for immunofluorescence were obtained from Invitrogen.
-
ZIKV strain ZIKA-SMGC-1 (GenBank accession number KX266255) was kindly provided by Dr. Ying Wu (Wuhan University). Cells were infected by ZIKA-SMGC- 1 in DMEM for 2 h and then replaced with DMEM supplemented with 2% FBS at 37 ℃ with 5% CO2. The recombinant lentiviral vector expressing GFP or NS5-GFP was packaged with psPAX2 and PMD2.G and transfected into HEK293T cells. Cells were infected by lentivirus for 8 h and then replaced fresh medium. Plasmids psPAX2 (#12260) and PMD2.G (#12259) were gifts from Didier Trono lab provided by Addgene.
-
Plasmids including pLV_Zika_Cv_FLAG (# 79628), pLV_Zika_PrM_FLAG (# 79631), pLV_Zika_NS3_FLAG (# 79635), pLV_Zika_FLAG_NS4A (# 79636), pLV_Zika_NS2B_FLAG (# 79637), pLV_Zika_NS4B_FLAG (# 79640), pLV_Zika_NS1_FLAG (Variant: W98G) (# 79641) and pLV_Zika_NS5_FLAG (# 79639) were gifts from Vaithi Arumugaswami lab provided by Addgene. And plasmids pLV_Zika_E_FLAG and pLV_Zika_NS2A_FLAG were constructed in the same vector using restriction enzyme digestion and ligation method with ZIKA-SMGC-1 genome as template. The ZIKV NS5 gene with a GFP tag was subcloned into a lentiviral vector under the EF1a promoter, which promoted protein expression in stem cells. The full length of ZIKV-NS5 and its truncation mutants with a FLAG tag, and the full-length p53 and its truncated proteins with HA tag at their Cterminus were constructed. The p53 with GST tag was constructed in plasmid pGEX 4 T-1 for expression and purification. All plasmid constructs were confirmed by sequencing.
-
The recombinant proteins with GST tag were expressed in Escherichia coli BL21(DE3) cells. Transformed bacteria were cultured in LB medium containing ampicillin (100 μg/mL) at 37 ℃ until the OD600 nm reached 0.6–0.8 and induced with 0.4 mmol/L isopropyl-b-D-thiogalactopyranoside (IPTG) at 16 ℃ for 20 h and then were suspended with lysis buffer (20 mmol/L tris–HCl, pH 7.0, 500 mmol/L NaCl, 5% glycerol, 1 mmol/L DTT, 1 × protease inhibitor cocktail) and incubated with lysozyme on ice for 30 min, and then sonicated on ice. The lysate was centrifuged at 10000 ×g for 10 min at 4 ℃. After centrifugation, the supernatant was incubated with GST agarose beads (GST beads) (GE healthcare) for 1 h at 4 ℃ with rotation and beads were eluted with elution buffer (20 mmol/L tris–HCl, 20 mmol/L reduced glutathione, 1 mmol/L DTT, 1 × protease inhibitor cocktail). The proteins were further concentrated by a membrane concentrator (Millipore).
-
HEK293T cells and hNPCs were seeded in chamber slides for transfection. Cells were fixed in 4% paraformaldehyde for 10 min, and permeabilized with 0.25% Triton X-100 for 10 min and then were blocked with 5% bovine serum albumin (BSA) for 30 min and incubated with primary antibodies overnight at 4 ℃. The coverslips were washed for 30 min in PBST and incubated with secondary antibodies for 30 min. Finally, cells were stained with 40, 6- diamidino-2-phenylindole (DAPI) to make the nuclei visible. For Hoechst-PI staining, hNPCs were prepared in cell culture plate and stained with Hoechst and PI according to the manufacturer's instruction of Hoechst 33342/PI Double Stain Kit (Solarbio). Immunofluorescence was visualized using a Zeiss LSM 800 Confocal microscope. And images were processed by ZEN (Zeiss) or Image J software.
-
The hNPCs expressing NS5 or control vector were collected 48 h post-infection by lentivirus. Annexin V-APC/PI apoptosis kit (Multi Science) was used to stain apoptotic cells and then cells were subjected to flow cytometry by using a flow cytometer (BD FACSAria III, BD).
-
Cells were lysed in RIPA buffer (50 mmol/L tris–HCl, 150 mmol/L NaCl, 1% TritonX-100, 1 mmol/L EDTA2Na, 0.5% SDS, 1% NP-40, 0.5% sodium deoxycholate and 1 × protease inhibitor cocktail) and clarified by centrifugation at 10000 ×g for 10 min at 4 ℃. Cell lysate samples mixed with SDS sample buffer were subjected to SDS-PAGE electrophoresis and transferred to PVDF membranes (Millipore). PVDF membranes were blocked in 5% bovine serum albumin and incubated with the corresponding antibodies.
-
Cells were collected 24 h post-transfection or 48 h post ZIKV infection and suspended in lysis buffer (50 mmol/L Tris–HCl pH 7.5, 150 mmol/L NaCl, 10% glycerol and 1 9 protease inhibitor cocktail) for 30 min on ice. After centrifugation, the supernatant was immunoprecipitated with anti-FLAG or anti-GFP agarose beads (Millipore) at 4 ℃ for 2 h with rotation. For denature-immunoprecipitation, the hNPCs were collected 48 h post-infection by lentivirus and suspended in lysis buffer (50 mmol/L Tris– HCl pH 7.5, 150 mmol/L NaCl, 10% glycerol and 1 × protease inhibitor cocktail) for 30 min on ice, and centrifugated at 10000 ×g for 10 min at 4 ℃. The supernatant with 10% SDS was boiled for 5 min and then was diluted with 1% SDS to incubate with anti-p53 antibody and protein A + G beads at 4 ℃ for 2 h with rotation. Beads were washed with lysis buffer for 5 times and were boiled in loading buffer. Samples were separated by SDS PAGE and then detected by immunoblotting as described above.
-
HEK293T cells were transfected with the construct expressing NS5 with a FLAG tag and lysed 24 h posttransfection. After centrifugation at 10000 ×g for 10 min at 4 ℃, the supernatant was collected to incubate with p53- GST or GST and GST beads for 2 h at 4 ℃ with rotation. Beads were washed 5 times with washing buffer (PBS with 1% Triton X-100) and then were boiled at 100 ℃ for 10 min for further detection.
-
Total RNA was extracted using Trizol reagent, and then 500 ng of RNA and the PrimeScriptTM RT Master Mix reverse transcription kit (Takara) were utilized for reverse transcription to synthesize cDNA. Quantitative PCR assays were performed with Power Up SYBR Green PCR Master Mix (Invitrogen) in ABI Q5 Detection 682 System (Applied Biosystems) using specific primers listed as follows: NOXAF (5′-GTGTGCTACTCAACTCAG-3′), NOXA-R (5′-ATT CCTCTCAATTACAATGC-3′); p21-F (5′-GAGACTCTCAGGGTCGAAAACG-3′), p21-R (5′-GGATTAGGGCTT CCTCTTGGA-3′); puma-F (5′-GACCTCAACGCACAGT ACGAG-3′), puma-R (5′-AGGAGTCCCATGATGAGATT GT-3′); p53-F (5′- GTGAGGGATGTTTGGGAGATG-3′), p53-R (5′- CCTGGTTAGTACGGTGAAGTG-3′).
-
The details of statistical analysis are described in each figure legend. Statistical analysis is performed on at least 3 biological replicates. The quantitative data were expressed as the mean ± SEM and all the statistical analyses were performed on a Graph-Pad Prism 8.3 software. The statistical significance between different groups was calculated by two-tailed unpaired Student' t-test. Difference was considered to be statistically significant when P < 0.05.
Cell Lines
Antibodies
Viruses and Infection
Plasmid Construction
Expression and Purification of Recombinant Proteins
Immunofluorescence Microscopy and Hoechst-PI Staining
Flow Cytometry
Immunoblotting
Co-Immunoprecipitation
GST Pull-Down
Quantitative Real-Time PCR
Statistical Analyses
-
The immortalized human neural progenitor cells (hNPC), ReNcell CX cells (Donato et al. 2007), which highly express the stem cell markers Nestin and Sox2 (Supplementary Fig. S1), were infected with ZIKV. Western-blot showed that ZIKV infection increased the cleavage of PARP (Fig. 1A), a well-known apoptosis marker, confirming that ZIKV infection could efficiently induce apoptosis of hNPCs. Meanwhile, the mRNA level of p53 target genes, NOXA, p21 and puma, was significantly upregulated by ZIKV infection (Fig. 1B) and the phosphorylation level of p53 at the serine 15 site was also apparently increased (Fig. 1C), indicating that p53 signaling pathway was activated in ZIKV-infected hNPCs. To investigate which of the ZIKV-encoded proteins was responsible for the activation of p53 signaling pathway, we expressed them in HEK293T cells, respectively, and measured the mRNA level of puma to observe the state of p53 pathway activation (Fig. 1D). The results showed that ZIKV-NS5 most significantly upregulated the transcription of puma, indicating its potential to activate the p53 pathway (Fig. 1E). Besides, ZIKV-NS2A also showed to increase the puma mRNA level, although to a lesser extent than NS5 (Fig. 1E). Thus, we focused to investigate the function and mechanism of ZIKVNS5 in the ZIKV-induced activation of p53 pathway.

Figure 1. ZIKV-NS5 activates p53 signaling pathway. A–C ZIKV infection caused the p53-mediated apoptosis of hNPCs. hNPCs were infected with ZIKV and cells were lysed 48 h post infection. The cleaved-PARP was detected by Western-blot (A), and gene transcription of p53-targeting genes was detected by RT-qPCR (B), and the expression of p53 downstream genes and phosphorylation of p53 were detected by Western-blot (C). D and E Effects of ZIKV-encoded proteins on the activation of p53 signaling pathway. The ten proteins of ZIKV were expressed in HEK293T cells, respectively, and the protein expression was detected by Western-blot (D), and the transcription of puma was detected by RT-qPCR (E). F and G NS5 activated p53 pathway in hNPCs. hNPCs were infected with lentivirus to express NS5 or vector, and cells were lysed 48 h post infection. The transcription of p53 and its targeting genes was detected by RTqPCR (F), and the protein expression and phosphorylation of p53 were detected by Western-blot (G). (B, E, F) n = 3 biological replicates. Data are Mean ± SEM. Two-tailed unpaired Student' s ttest was used for statistical analysis. *P < 0.05, **P < 0.01, ***P < 0.001, ns. = not significant.
NS5 protein is the RNA-dependent RNA polymerase (RdRp) of ZIKV, which is responsible for the replication of the viral genome in the cytoplasm. However, it is predominantly localized in the nucleus, implying that it may perform additional functions (Hou et al. 2017). When expressed in hNPCs, ZIKV-NS5 significantly promoted the transcription of p53 downstream genes, including p21 and puma, while had minimal effects on the mRNA level of p53 itself (Fig. 1F). In consistency with that of the viral infection (Fig. 1C), ZIKVNS5 expression also increased the protein level and the S15- phosphorylation of p53 (Fig. 1G). Together, these data indicated that among ZIKV-encoded proteins, ZIKV-NS5 plays a crucial role in the activation of p53 signaling pathway.
-
Subsequently, the immunofluorescence staining showed that expression of ZIKV-NS5 significantly induced the accumulation of p53 in the nuclei of both HEK293T cells and hNPCs (Fig. 2A and 2B). Given that ZIKV-NS5 did not affect mRNA level of p53 while increased its protein level, we speculated that p53 was regulated at post-transcriptional stages. Therefore, we treated the hNPCs with cycloheximide (CHX), an inhibitor of protein translation. The results showed that the protein level of p53 dramatically decreased and almost disappeared 6 h after the treatment in the control group (vector), while the decrease rate of p53 level was much slower in ZIKV NS5-expressing cells and a portion of p53 protein remained after 8 h of treatment (Fig. 2C and 2D). This indicated that the existence of ZIKV-NS5 increased the half-life of p53. Furthermore, to investigate how NS5 stabilizes p53, cells expressing NS5 or vector were treated with etoposide and then collected for immunoprecipitation with anti-p53 antibody. The results showed that the expression of NS5 protein inhibited the p53-MDM2 interaction (Fig. 2E). Subsequently, we found that the expression of NS5 protein repressed the ubiquitination of p53 (Fig. 2F). Altogether, these results suggested that ZIKV-NS5 expression could stabilize p53 in the nucleus and consequently promote the expression of its downstream genes.

Figure 2. NS5 protein promotes the accumulation of p53 in nuclei and prolongs its half-life. A and B NS5 promoted the accumulation of p53 in nuclei. NS5-expressing hNPCs and HEK293T cells were performed immunofluorescence with anti-p53 antibody and the nuclei was stained with DAPI (A). NS5 (green) and p53 (red) and DAPI (blue). Scale bar, 5 μm. B The fluorescence intensities of p53 in NS5 negative hNPCs (n = 108) or positive cells (n = 58) (left panel) and in NS5 negative HEK293T cells (n = 111) or positive cells (n = 60) (right panel) were analyzed by image J software. Data are Mean ± SEM. Two-tailed unpaired Student' s t-test was used for statistical analysis. ****P < 0.0001. C and D NS5 increased the half-life of p53. hNPCs were infected by lentivirus to express NS5 or vector and were treated with 50 μg/mL cycloheximide (CHX) for the indicated times. The protein level of p53 was detected by Western-blot (C) and the immunoblot signals of p53 were analyzed by image J software and normalized to the GAPDH signal and then plotted against the signal obtained at 0 h of cycloheximide treatment (D). n = 3 biological replicates. Data are Mean ± SEM. Two-tailed unpaired Student' s ttest was used for statistical analysis. *P < 0.05, **P < 0.01, ***P < 0.001, ns. = not significant. E NS5 inhibited the p53- MDM2 binding. Cells expressing NS5 or vector, were treated with 50 μmol/L etoposide for 24 h and collected for immunoprecipitation with anti-p53 antibody. The protein interaction was detected by Western-blot. F NS5 protein repressed the ubiquitination of p53. Cells expressing NS5 or vector were treated with MG132 for 12 h and the not treated group was set as the control, and then were collected for denature-immunoprecipitation with anti-p53 antibody. The ubiquitination of p53 was detected by Western-blot.
-
To further investigate the detailed relationship between ZIKV-NS5 and p53 pathway, we detected the interacting partners of ZIKV-NS5 by co-immunoprecipitation (Co-IP) followed with mass spectrometry (MS) analysis. Intriguingly, the results showed that p53 itself was captured by ZIKV-NS5 (Fig. 3A and 3B). Further Co-IP experiments confirmed that NS5 and p53 could be co-immunoprecipitated by each other (Fig. 3C and 3D). Moreover, in ZIKVinfected hNPCs, NS5 could be immunoprecipitated by endogenous p53 (Fig. 3E). In addition, NS5 and p53 also showed co-distribution in 10%–40% (v/v) sucrose gradient centrifugation assay in both HEK293T cells and hNPCs lysates (Fig. 3F). To further assess this interaction, GST pull-down assay was performed, and the result showed that ZIKV-NS5 could be pulled down by the recombinant p53- GST but not GST (Fig. 3G). Collectively, these data indicate that ZIKV-NS5 could directly interact with p53.
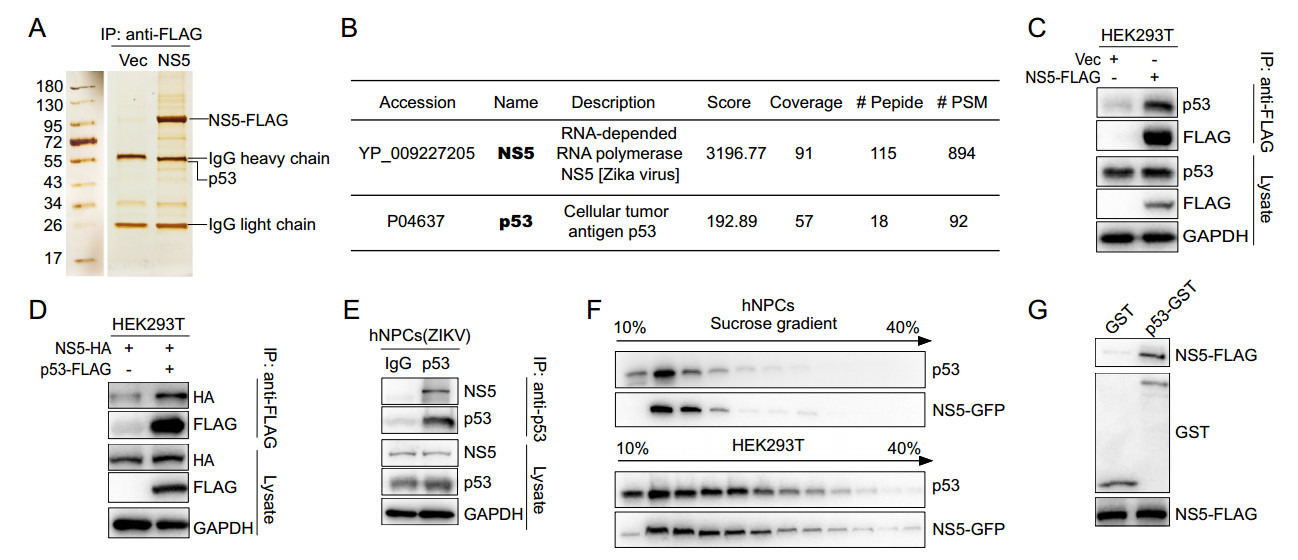
Figure 3. NS5 interacts with p53. A and B The p53 protein was immunoprecipitated by NS5. HEK293T cells, transfected with plasmid encoding NS5-FLAG or vector, were lysed and immunoprecipitated with anti-FLAG beads. The elution from anti-FLAG beads were separated by SDS-PAGE and stained by sliver-staining (A). The whole lanes of NS5 or vector were analyzed by mass spectrometry (MS) assay and p53 were detected (B). C and D NS5 and p53 was immunoprecipitated by each other. HEK293T cells were co-transfected with the indicated plasmids and then co-immunoprecipitation assay was performed with anti-FLAG beads. The endogenous p53 (C) and NS5-HA (D) were detected by Western-blot. E The ZIKVencoded NS5 was immunoprecipitated by p53. hNPCs infected by ZIKV were lysed and immunoprecipitated with anti-p53 antibody. The NS5 protein was detected by Western-blot. F NS5 and p53 codistributed in sucrose gradient centrifugation experiment. hNPCs and HEK293T cells that expressed NS5 were lysed and fractionated on 10%–40% (v/v) of sucrose gradients and detected by Western-blot. G NS5 interacted with p53 in vitro with GST pull-down assay. The p53-GST and GST proteins were expressed in Escherichia coli BL21(D3) cells and purified to incubate with NS5-expressing HEK293T cells lysate, and then bound with GST beads. The protein interactions were detected by Western-blot.
-
To determine the precise regions responsible for the NS5-p53 interaction, a series of truncations were constructed (Fig. 4A and 4B). NS5 consists of an N-terminal methyltransferase (MTase, residues 1–264) domain and a C-terminal RNAdependent RNA polymerase (RdRp, residues 265–903) domain (Fig. 4A). Truncated proteins of NS5 were expressed in HEK293T cells, respectively, and through Co-IP assays, the MTase domain of NS5 was detected to mediate the interaction of NS5 and p53 (Fig. 4C). The p53 protein has a transactivation domain (TAD, residues 1–97) at the N-terminus, and followed a core DNA binding domain (DBD, residues 102–292), and a C-terminal domain (CTD, residues 293–393) (Jenkins et al. 2012) (Fig. 4B). Similarly, truncated proteins of p53 were co-expressed with NS5 in HEK293T cells and the co-immunoprecipitation results showed that NS5 could bound with the CTD domain of p53 (Fig. 4D). Thus, the MTase domain of NS5 and the CTD domain of p53 were directly involved in their interaction of NS5 and p53.
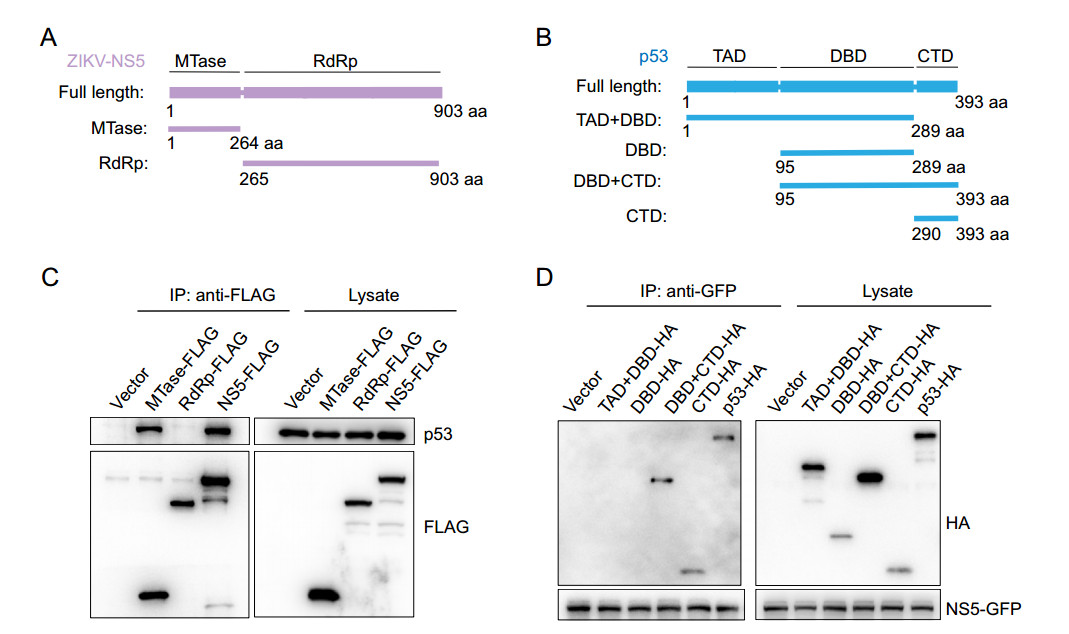
Figure 4. MTase domain of NS5 and CTD of p53 mediates the NS5-p53 interaction. (A and B) Schematic diagram of truncation mutants of ZIKV-NS5 (A), and p53 (B). C and D MTase domain of ZIKV-NS5 and the C-terminal domain (CTD) of p53 mediated their interaction. HEK293T cells transfected plasmids that expressing ZIKV-NS5, MTase, RdRp or vector (C) and cells co-transfected ZIKV NS5- expressing plasmid with plasmids expressing p53, TAD + DBD, DBD, DBD + CTD, CTD or vector (D), were lysed and immunoprecipitated. The protein interactions were detected by Western-blot.
-
Given that ZIKV-NS5 could activate p53 signaling pathway similar to the virus infection, we wonder whether it could also cause apoptosis in hNPCs. We found that morphological changes of hNPCs apparently took place (Fig. 5A) and the fluorescence signal of dead cells was significantly increased in NS5-expressing hNPCs (Fig. 5B). Furthermore, Western-blot analysis showed that NS5 expression increased the level of cleaved PARP (Fig. 5C). The flow cytometry assay indicated that the percentage of apoptotic cells was significantly elevated by ZIKV-NS5 (Fig. 5D and 5E). To prove the role of p53 in NS5-induced cell apoptosis, we depleted the expression of p53 in hNPCs by specific siRNA and further expressed ZIKV-NS5 in these cells by lentivirus infection. And we found that the siRNA successfully depleted p53 expression and ZIKV-NS5 failed to induce the cleavage of PARP protein in hNPCs (Fig. 5F), implying that cell apoptosis was thereby inhibited, and ZIKV-NS5-induced apoptosis was p53-dependent. Altogether, these results suggested that, consistent with ZIKV infection, ZIKV-NS5 could induce apoptotic cell death of hNPCs through regulating p53 activity.
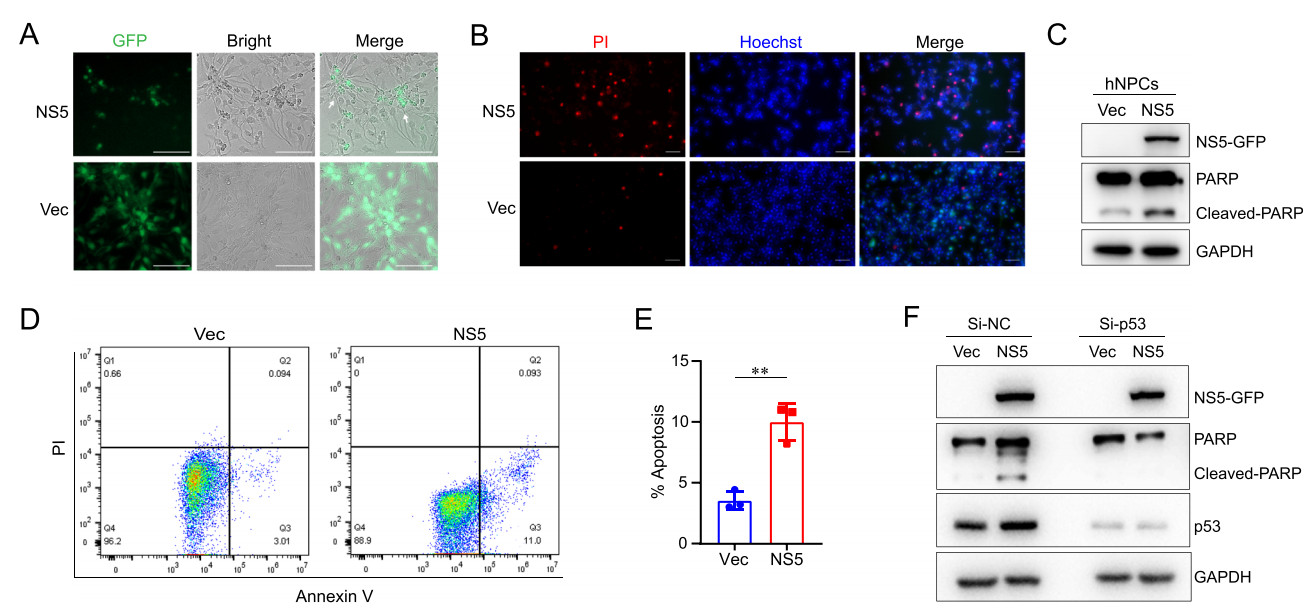
Figure 5. ZIKV-NS5 induces apoptosis of human neural progenitor cells. A and B NS5 expression caused obvious morphological changes and cell death in hNPCs. hNPCs were infected with lentivirus to express NS5-GFP or vector, and the cells were observed by fluorescence microscope (A). Scale bar, 100 μm. And hNPCs were stained with PI and Hoechst double staining (B). PI (red) and Hoechst (blue). Scale bar, 50 μm. C–E ZIKV-NS5 induced the apoptosis of hNPCs. hNPCs expressing NS5 or vector were lysed to detect the cleaved-PARP (C) and the proportions of apoptotic cells were analyzed by flow cytometer assays (D and E). n = 3 biological replicates. Data are Mean ± SEM. Two-tailed unpaired Student' s t-test was used for statistical analysis. **P < 0.01. F Knocking down of p53 inhibited NS5-induced cell apoptosis in hNPCs. The hNPCs were transfected with siRNA that specifically targeting p53, and then were infected by lentivirus to express NS5 or vector and the cells were collected.
-
Since the out break in South America in 2015, the ZIKV isolations could cause microcephaly in neonates born to mothers with intrauterine infections. The apoptosis-induced cell death of NPCs mediated by p53 activation has been reported to play a pivotal role in ZIKV caused microcephaly (Ghouzzi et al. 2016; de Sousa et al. 2018). In this study, we investigated the effects of ZIKV-encoded proteins on the p53 signaling pathway and found that ZIKV-NS5 significantly promoted the p53 stability through repressing the p53- MDM2 binding and subsequently inhibiting the ubiquitination of p53, and activated the transcription of p53 downstream genes. After viral infection, ZIKV-NS5 was localized in the nuclei and directly interacted with p53 through its MTase domain. This interaction stabilized p53 from degradation and then activated p53-mediated apoptosis of hNPCs.
NS5 protein is the RNA-dependent RNA polymerase (RdRp) of flaviviruses, which is responsible for the replication of viral genome in the cytoplasm (Abrams et al.2017). However, consistent with previous reports (Hou et al. 2017), we observed that ZIKV-NS5 was predominantly localized in the nucleus (Fig. 2A). Such a localization implies that, apart from its role in viral RNA replication, ZIKV-NS5 may perform additional functions in nuclei. Indeed, ZIKV-NS5 has been reported to interact with splicing factor SC35 and the RNA polymerase-associated factor 1 complex (Paf1C) in the nucleus to regulate the RNA splicing and transcriptional elongation processes of the host cells (Hou et al. 2017; Shah et al. 2018). In the present work, we demonstrated that the nuclear ZIKV-NS5 associates with p53 protein and then prolonged its half-life, leading to the upregulation of the p53 downstream genes. This is the first study, to the best of our knowledge, to report the direct interaction between p53 and RdRP of a flavivirus. Similar to ZIKV-NS5, the NS5 proteins of Dengue virus (DENV) and yellow fever virus (YFV) is also documented to localize in the host cell nucleus (Tay et al.2013; Laurent-Rolle et al. 2014). DENV- and ZIKV-NS5 showed high similarity in amino acid sequence (Supplementary Fig. S2A), and intriguingly, we also found that DENV-NS5 protein could interact with p53 (Supplementary Fig. S2B), but ZIKV and DENV show different tropism to host cells and NPCs are not the target cells of DENV (Brault et al. 2016; Muffat et al. 2018). It will be meaningful to evaluate whether DENV-NS5 would influence the fate of its natural host cells in future work.
Abnormal activation of p53 signaling pathway is responsible for hNPC apoptosis induced by various factors and microcephaly during fetal brain development (Toki et al.2018). Ablation of p53 can extensively rescue the microcephaly phenotype caused by genetic mutations (Marjanovic et al. 2015; Breuss et al. 2016; Bianchi et al. 2017). Our results proposed a causal link between NS5 and p53-dependent apoptosis of the hNPCs. Further efforts with ZIKV-NS5 transgenic animal model are needed in the future to investigate whether the expression of ZIKV-NS5 alone can recapitulate the microcephaly phenotype of ZIKV infection and whether p53 deficiency could rescue such a phenotype. Notably, NS5 is not the only ZIKV protein that can affect p53 signaling pathway. We found that NS2A also showed to upregulate the expression of puma, although to a lesser extent than NS5 (Fig. 1E). However, during the preparation of this manuscript, another study reported that NS2A could bind to p53 but dampen p53 transcriptional activity (Alzhanova et al. 2021). This discrepancy may be caused by the different systems were used in the two studies: we detected the expression level of endogenous p53 target genes in HEK293T cells without drug stimulating, while Alzhanova et al. (2021) used p53-luc reporter system in U2OS cell with etoposide-induced DNA damage condition. Further studies are needed to verify the exact role of ZIKV-NS2A in the virus-induced p53 pathway activation in hNPCs and even in animal models. Moreover, ZIKV capsid protein has also been reported to stabilize p53 through interacting with and blocking the E3 activity of MDM2 (Teng et al. 2017). Taken together, ZIKV may have developed various mechanisms to modulate the activity of p53 signaling pathway in host cells, and our findings reveal that ZIKV-NS5 is a previously undiscovered key player.
In summary, this study reveals that ZIKV-NS5 could directly activate p53-mediated apoptosis of hNPCs, highlighting its potential contribution to the microcephaly caused by ZIKV infection. These findings further expanded our knowledge about the interaction between ZIKV and its host cells and provide a potential therapeutic target for the treatment of the virus-induced neurodevelopmental defects.
ZIKV-NS5 Activates p53 Signaling Pathway
ZIKV-NS5 Promotes the Accumulation of p53 in the Nuclei and Its Protein Stability
ZIKV-NS5 Interacts with p53
The MTase Domain of NS5 and the CTD Domain of p53 are Involved in the NS5-p53 Interaction
ZIKV-NS5 Induces Apoptosis of Human Neural Progenitor Cells
Discussion
-
The work is supported by the National Natural Science Foundation of China [NSFC Grant#81620108020 and #32041002, to D.G.; Grant #31800151, to J.W.], Guangdong Zhujiang Talents Program (to D.G.), Shenzhen Science and Technology Program [Grant #KQTD20180411143323605 and # JSGG20200225150431472 to D.G.] National Ten-thousand Talents Program (to D.G.), Guangdong Province "Pearl River Talent Plan" Innovation and Entrepreneurship Team Project (Grant #2019ZT08Y464 to Li, C.M). We are grateful to Dr. Ying Wu (Wuhan University) for sharing the ZIKV SMGC-1(GenBank accession number: KX266255) and to Prof. Mengfeng Li (Sun Yat-Sen University) for sharing the human neural progenitor cells and to assistants in the two laboratories for sharing the culturing experience.
-
PL, JW, DG and CL contributed to the conception of the study. PL and HJ performed the experiments and analyzed the data. PL and HJ prepared the figures and wrote the manuscript. WZ, YZ and MH, LX and JC assisted with the experiments. JW, HP, DG and CL edited the manuscript and oversaw all study design and data analysis. All authors read and approved the final manuscript.
-
The authors declare that they have no conflict of interest.
-
This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: