









HTML
-
The Enterobacteriaceae are a class of gram-negative bacteria. Within this family, all bacterial species have been implicated in bloodstream, cholangitis, peritonitis and other intra-abdominal infections(Paterson, 2006). Additionally, Escherichia coli often causes urinary tract infections; Klebsiella pneumoniae has been regarded as an important cause of pneumonia; Salmonella enterica produces gastroenteritis, and subsequently invasive infection in some patient; and Shigella strains could cause shigellosis, which often occurs in epidemic form and causes considerable morbidity and mortality(Donnenberg, 2002; Paterson, 2006; Phalipon and Sansonetti, 2007; Verma et al., 2010). Unfortunately, emerging drug resistance is a serious problem in handling Enterobacteriaceae infections, and threatens to create pan-resistant species(Falagas et al., 2010). There is a pressing need to find alternative ways to control Enterobacteriaceae infections, and one of the efficient methods is bacteriophage therapy.
Bacteriophage therapy has a long history, which originated in the early 20th century. Since the discovery of penicillin and other antibiotics, with their easy use and notable effects in treating microbial related diseases, little attention has been paid to bacteriophage therapy for nearly half a century(Burrowes et al., 2011). However, due to the abuse of antibiotics, a greater number of pathogenic bacteria becoming resistant to drugs, resulting in great threat to human health(Merril et al., 2003). In China, the situation is more serious. Susceptibility rates of clinically isolated Enterobacteriaceae declined by about 30% against the new generations of cephalosporins from 2002 to 2009; meanwhile, there was a rapid increase in the occurrence of extended-spectrum β-lactamases, especially for the E. coli strains(from 20.8% in 2002 to 64.9% in 2009)(Yang et al., 2010). Three reasons lead to this situation: the first was the abuse of antibiotics; the second was the lack of antibiotics with new antibacterial mechanisms; and the third was the investment atrophy for the d evelopment of new antibiotics(Sulakvelidze et al., 2001). More seriously, t he rate of development of new antibiotics is slower than t he rate of the appearance of antibiotic resistance; therefore, the prospect of new kinds of drugs for clinical use is not optimistic. These realities have stimulated researchers in the study of bacteriophage therapy, which has met with great progress(Sulakvelidze et al., 2001).
-
The history of phage therapy started with phage discovery. Briefly, Hankin first reported the presence of an antibacterial activity against Vibrio cholera in 1896(Hankin E, 1896). Two years later, Gamaleya discovered the same phenomenon when working with Bacillussubtilis(Samsygina and Boni, 1984; Sulakvelidze et al., 2001). Twort first reported the discovery of plaque in plates during the culture of Staphylococcus aureus, and explained it as a viral infection in 1915(Twort, 1915). In 1917, d'Herelle proved the phages therapy(d'Herelle, 1917). He used phages to treat Shigell astrains isolated from patients with dysentery after which small and clear areas appeared on the agar plates. He proposed that the phenomenon was due to parasitizing bacteria caused by virus infection. The name "bacteriophage" was also proposed by d'Herelle; it was derived from the words "bacteria" and "phagein"(dHerelle, 1917; Summers, 1999). Another important contribution by d'Herelle was that, he firmly promoted the idea that phages were live viruses, but not "enzymes" as many researchers thought in his era. The research and application of bacteriophages was started thereafter. In 1919, a boy with dysentery in Paris was treated with d'Herelle's anti-dysentery phage and recovered within a few days. Soon after, three patients with bacterial dysentery were also recovered after being treated with d'Herelle's anti-dysentery phage, which confirmed the efficacy of phage therapy. Unfortunately these studies were not reported. T he first report of phage therapy came in 1921 when Bruynoghe and Maisin used bacteriophages to treat staphylococcal skin disease(Bruynoghe and Maisin, 1921; Payne et al., 2000). In 1932, East European scientists identified the scientific dosage of phage based on abundant animal and human tests. Thereafter, several companies, such as the ParkeDavis Company and Eli Lilly and Company, began active commercial production of phages against various bacterial pathogens. In China, phage therapy was started in 1955 when Si et al.successfully used bacteriophages to treat Shigella dysenteriae(Si, 1955). However, in these early studies, the phages used had a narrow host range, low purity and instability, which limited the application of phage therapy(Qian et al., 2007).
-
Bacteriophage therapy is the therapeutic use of phages to split pathogenic bacteria. After adsorption to bacteria, phages start the process of bacteriolysis. According to the requirement of e ndolysin or not, host cell lysis mechanisms can be divided into two basic modes. One mode depends on the lysozymes produced by phages with dsDNA such as phages K and T4 to split bacteria; the other mode does not require the lysozymes such as phage MS2 with ssRNA and phage φX174 with ssDNA(Young, 1992). Hense, there are two basic types of bacterial lytic mechanisms.
-
This type of bacteriophages lacks the genes encoding lysozymes, and they split host strains by synthesizing proteins to inhibit the host cell wall biosynthesis, leading to lysis of host cells during growth. E. coli ssRNA phage Qβencodes a protein A2 and binds to protein MurA, an enzyme that catalyzes the first step in cell wall synthesis, preventing catalysis by occluding phosphoenolpyruvate from accessing the active site(Reed et al., 2012). The ssDNA phage φX174 encodes a membrane protein E, which inhibits the enzyme activity of MraY and results in host cell lysis(Tanaka and Clemons, 2012). MraY catalyzes the first membrane-localized step for the synthesis of peptidoglycan precursor(Tanaka and Clemons, 2012). During this type of lysis, the host cell wall forms small lesions, and leaves the ghost or large cell debris at last(Young, 1992).
-
Other than the lysozyme independence of some bacteriophages, most bacteriophages have dsDNA encode lysozymes to crack the host cells. The dsDNA bacteriophages belong to the order Caudovirales, and account for about 95% of all the bacteriophages discovered(McAuliffe et al., 2007). This type of phage splits the host cells by expressing two types of lysozymes, holins and lysins.
Bacteriophage-encoded holins are a diverse group of membrane proteins. Holi ns control the transmembrane or activity of phage-encoded endolysins to degrade the host cell walls, and thereby initiate the bacteriolytic process as the "lysis clock"(Young and Bläsi, 1995). Based on structural difference, Young et al. classified phage holin s into three groups, class Ⅰ, Ⅱ and Ⅲ(Figure 1)(Young and Bläsi, 1995).
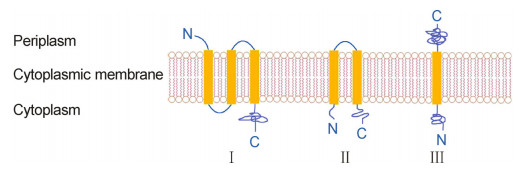
Figure 1. Topology structure of phage holins. Phage holins can be divided into three groups, classes Ⅰ, Ⅱ and Ⅲ.
The typical class Ⅰ holin is S105 coded by the S gene of phage λ, which contains 105 amino acids, and forms three transmembrane domains(TMDs)(Gründling et al., 2000). The zwitter ionic and non-ionic detergents permit S105 to form oligomers, and the oligomers form ringshaped structures(Savva et al., 2008). The ring has an external diameter of 23 nm, an inner diameter of 9 nm for the upper ring and 8 nm for the lower ring(Savva et al., 2008). The 8 nm inner diameter of the ring is large enough for the movement of endolys in R of phage λ to the periplasm where it attacks the host cell wall. Studies also show that the amnio acid mutation of S105 could affect the lysis time(Gründling et al., 2000).
The class Ⅱ holins have two TMDs, which usually have a length of 65 to 95 amino acids(Shi et al., 2012). The S21 protein from lambdoid phage 21 is a holin of this type(Pang et al., 2009). In the early stage of cytoplasmic membrane damage, both the N-and the C-terminus of S21 are in the cytoplasm, and form small membrane holes that depolarize the membrane, thus class Ⅱ holins are also considered as pinholins, as opposed to large-holeforming holins like S105. The lysins of this type of phage are exported by the sec system of host cells and accumulate in the inactive form tethered to the membrane by an N-terminal SAR("signal-anchor-release")sequence in the periplasm. The depolarization by S21 leads to the release of the lysin from the biolayer and splits cell wall. During this process, the holin triggers the depolarization of the cell membrane, and leads to the release of the SAR lysin from the bilayer; in this way it is thought to impose the timing on the lytic event(Pang et al., 2009; Xu et al., 2004).
The class Ⅲ holin has only one TMD, which is highly hydrophilic. A representative holin is T protein from phage T4(Ramanculov and Young, 2001). The T protein of bacteriophage T4, as other holins, has the ability to cause the formation of a lethal membrane lesion, and allows the phage lysin to target the cell wall. The N-terminus of T is in the cytoplasm and acts as membrane lesion; the C-terminus is in the periplasm and functions as the regulator. Moreover, T acts in the vegetative cycle like other holins at an accurate programmed time(Ramanculov and Young, 2001).
Lysins, also known as endolysins, muralysins, muramidases or virolysins, are highly evolved enzymes to digest the bacterial cell wall for the release of phage progeny during the final stage of bacteriophage lytic cycle(Fischetti, 2008). Functionally, four types of phage lysins can be classified: endo-β-N-acetylglucosaminidase, N-acetylmuramidase, endopeptidase and N-acetylmuramoyl-L-alanine amidase(Loessner, 2005; Pastagia et al., 2013). The targets of these lysins on host cell wall are shown in Figure 2. Due to the high effectiveness and specificity to drugs, the lysins have been applied as antibacterial agents(Fischetti, 2008). Lysins must perform two basic functions: substrate recognition and enzymic hydrolysis(Díaz et al., 1990). Generally, the N-terminal domain cleaves specific peptidoglycan bonds, such as endo-β-N-acetylglucosaminidase and N-acetylmuramidases which hydrolyse glycosidic bonds in the glycan strand, endopeptidases cleave the peptide bonds of the cross-bridge, and N-acetylmuramoyl-Lalanine amidases cleave the amide bond connecting the glycan moiety and the stem peptide(Pastagia et al., 2013).
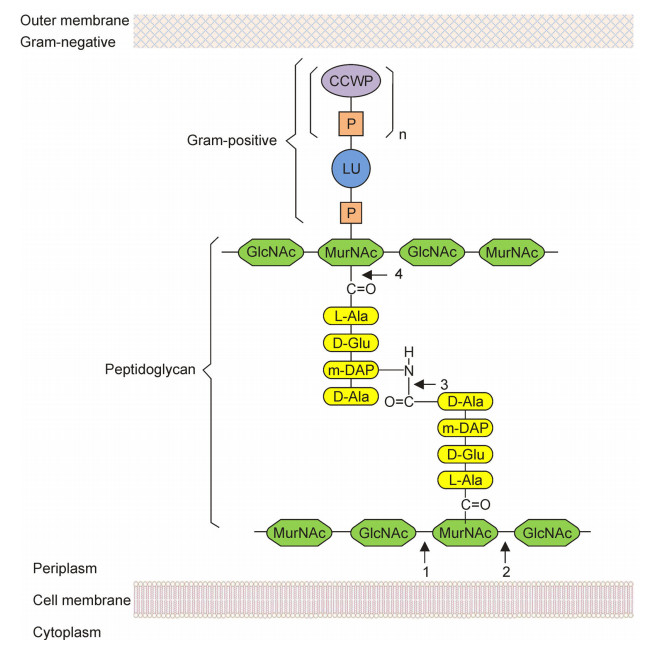
Figure 2. Lysins targeted on the peptidoglycan of bacterial cell wall (Adapted from Loessner, 2005). CCWP: carbohydrate cell wall polymer; P: phosphate group; LU: linkage unit; m-DAP: meso-diaminopimelic acid; GlcNAc: N-acetylglucosamine; MurNAc: N-acetylmuramic acid. 1: Endo-β-N-acetylglucosaminidase; 2: N-acetylmuramidase; 3: Endopeptidase; 4: N-acetylmuramoyl-L-alanine amidase.
We can see the difference between lysozyme independent and dependent lysis systems. Phages with lysozyme independent lysis system produce no holins or lysins. These phages encode proteins or peptides to inactivate the enzymes of synthesis pathways of the host cell wall, thus they function only on the cells in the growth stage; bacterial ghosts or large debris are left after cell lysis, and this mechanism often exists in the ssRNA or ssDNA phages. On the contrary, phages with lysozyme dependent lysis system use a combination of holins and lysins to split the host cells at any growth stage, the host cells are thoroughly lysed; this system is often found in dsDNA phages.
As an alternative to antibiotics, bacteriophages have a long history of treatment of a variety of bacterial diseases(Sulakvelidze et al., 2001). The following is a summarization of studies regarding phage therapy and Enterobacteriaceae.
-
It is estimated by the World Health Organization that about 5 million children die each year as a consequence of acute diarrhea(Snyder and Merson, 1982). One third cases of childhood diarrhea in developing countries are caused by E. coli(Albert et al., 1995). Due to its malleable genetic characteristic, E. coli has one of the widest spectra of disease of any bacterial species(Donnenberg, 2002). The E. coli O157 that recently emerged as a major food pathogen is a live example(Sharma et al., 2009). Furthermore, there is a lack of effective treatment for E. coli infections. Oral rehydration acts as the mainstay of treatment(Bhan et al., 1994). Countless lives have been saved by this simple and inexpensive measure. However, it does not affect the natural course of disease or the intrinsic activity of anti-bacteria. The use of antibiotics is of doubtful value since antibiotic resistance is widespread(Savarino et al., 2002).
Bacteriophages have been used to treat E. coli infections. Smith et al. used a mixture of phages to treat diarrhea in calves, piglets and lambs(Smith and Huggins, 1983). Similar results were achieved among these different animals; the diarrhea caused by the enteropathogenic E. coli strain was much reduced in vivo, and treatment had an ameliorating effect on the course of the disease. Recently, studies have also evaluated the safety of phage treatment through murine and human tests(Denou et al., 2009; Sarker et al., 2012). Denou et al. used a T4 coliphage to treat E. coli diarrhea by a combination of in vitro and in vivo tests(Denou et al., 2009). They selected phages based on genome sequencing and bioinformatic analysis, and found that the coliphage had a significant curative effect and no negative impact or anti-T4 antibodies were present after one month of treatment. Sarker et al. selected 9 phages without horizontal gene transfer and undesired genes from 99 T4-like coliphages based on genome sequencing to produce the cocktail, and gave it to 15 healthy adults from Bangladesh; no phage amplification or side effects were observed, which might indicate the safety of phage therapy(Sarker et al., 2012).
-
S. enterica is the Enterobacteriaceae family bacteria which can cause human salmonellosis(Paterson, 2006). S. enterica can live in the gastrointestinal tracts of birds and mammals, which could reptile and survive for a long time in environments such as water, soil and kinds of foods(Capparelli et al., 2010). Ingestion of food with animal feces is the cause of most human cases of salmonellosis. Clinical manifestations of human salmonellosis have a wide range, from subclinical gastroenteritis to severe bacteremia and meningitis, as well as other forms of extraintestinal infections(Angulo et al., 2000). In fact, more than 2300 serovars of S. enterica are now known, and display great differences in virulence(Heithoff et al., 2008). Because of the widespread presence of antibiotic resistance among S. enterica isolates, the only available antibiotics are the fluoroquinolone and third-generation cephalosporins(Angulo et al., 2000), hence there is a need for antimicrobial alternatives other than antibiotics.
The phage was firstly exploited for classifying S. enterica bacteria by its specificity for bacterial species or serovars(Anderson et al., 1977). Recently, phages have been used for the control of S. enterica contamination(Atterbury et al., 2007; Leverentz et al., 2001; Wall et al., 2010). Leverentz et al. used Salmonella-specific lytic phages to reduce the colony numbers in experimentally contaminated fresh-cut melons and apples(Leverentz et al., 2001). They found that the phages could significantly reduce Salmonella populations of melons at different temperatures, but did not work well on apples due to the low pH, which might affect the phage activity. Atterbury et al. tried to reduce Salmonella colonization of broiler chickens through the use of host-specific phages(Atterbury et al., 2007). Three broad host range phages were selected from 232 Salmonella phages. Two of the three chosen phages showed an obvious reduction of S. enterica serotype enteritidis cecal colonization. Their studies also indicated the key factors in the successful phage-mediated control of salmonellas were appropriate phages and optimization of both the timing and method of phage delivery(Atterbury et al., 2007). Wall et al. used a phage cocktail to treat small and market-weight pigs, and the results showed that the group of pigs with the phage cocktail had a significantly reduced ileal Salmonella concentrations as well as cecal Salmonella concentrations (Wall et al., 2010).
-
K. pneumoniaeis a gram-negative opportunistic pathogen, which is often associated with pneumonia, urinary tract, bloodstream, and intra-abdominal infections(Verma et al., 2010).K. pneumoniae caused bacteremia usually leads to significant morbidity and mortality among the general population(Tsay et al., 2002). More seriously, due to the prevalence of multidrug-resistant K. pneumoniae strains, treatment of these infections becomes ever more difficult(Verma et al., 2009).
Virulent phages specific to K. pneumoniae cells have been studied to control the infection caused by K. pneumoniae(Verma et al., 2009; Chhibber et al., 2008). Malik et al. used bacteriophage KØ1 to treat third-degree burn wounds of mice administered with a fatal dose of K. pneumoniae(Malik and Chhibber, 2009). After treatment, a substantial bacterial load decrease was noted in the blood, peritoneal lavage, and lung tissue of mice compared with those of the control groups. The decrease in microbial count was evident via the subcutaneous or intraperitoneal bacteriophage therapy. Gu et al. established a "step-by-step" approach to take advantage of the occurrence of phage-resistant bacteria variants(Gu et al., 2012). A phage cocktail consisted of three phages was established for K. pneumoniae by this method. The phage cocktail significantly reduced the mutation frequency of K. pneumoniae compared with any single phage, and effectively rescued K. pneumoniae bacteremia. Besides, the minimal protective dose of the phage cocktail was significantly smaller than that of single monophage to protect bacteremic mice from lethal K. pneumoniae K7 infection. Hung et al. treated K. pneumoniae -induced liver infection by using an isolated phage φNK5(Hung et al., 2011). The results indicated that a single dose of lower than 2 × 108 PFU phages was effective. The mice, via intraperitoneal or intragastric treatment, showed an elimination of K. pneumoniae from both blood and liver tissues compared with those of the control groups. This work suggested the low dose of φNK5 is an efficient therapeutic agent against K. pneumoniae -induced liver infection. Chhibber et al. used phage SS as the agent against an experimental model of K. pneumoniae -mediated lobar pneumonia in mice(Chhibber et al., 2008). A single intraperitoneal injection of 1010 PFU/mL phage administered immediately after intranasal challenge of 108 CFU/mL of K. pneumoniae B5055 was sufficient to rescue K. pneumoniae-mediated respiratory infections. A significant protection was observed in infected mice by administration of the phage preparation three hours prior to intranasal bacterial challenge. However, the phage treatment was ineffective even six-hour delay of phage administration following the induction of infection. Therefore, the results of this study suggested that the timing of initial phage therapy after initiation of infection significantly contributed to the success of treatment. Although few phage therapies on human K. pneumonia infection have been reported, the studies suggest that bacteriophages or bacteriophage cocktails have the poten-tial to modulate the infection caused by K. pneumonia.
-
Shigella is a kind of small, unencapsulated, non-motile gram-negative rod bacteria that caused shigellosis in humans. There are four species of Shigella pathogenic to human: S. dysenteriae, S. boydii, S. sonnei and S. flexneri(Subekti et al., 2001). Shigellosis is a major public-health problem in many developing countries, and causes considerable morbidity and mortality(Niyogi 2005; Phalipon and Sansonetti, 2007). Shigellosis causes an estimated 120 million cases and 1.1 million deaths worldwide annually(Niyogi, 2005). Shigella contamination occurs mainly through the fecal-oral route, with fomites, food, water, insects or direct person to person contact, and the infectious dose is as little as 100 bacterial cells(Phalipon and Sansonetti, 2007). In addition, secondary transmission through environmental sources cannot be ignored(Levine and Levine, 1991).
Bacteriophage treatment of Shigella originated in 1917, while d'Herelle first used phages to split Shigella strains isolated from several soldier patients with hemorrhagic dysentery(d'Herelle, 1917; Summers, 1999). One of the most extensive studies was conducted to evaluate the utility of therapeutic phages for prophylaxis of bacterial dysentery during 1963 and 1964 in Tbilisi, Georgia(Babalova et al., 1968). In total, 30, 769 children between 6 months to 7 years of age were included in the study. Of these children, 17, 044 on one side of the street were given Shigella phages orally, and the remainder on the other side of the street were not treated with phages. The final results showed that the incidence of dysentery of the placebo group was 3.8-fold higher than that of the phage-treated group, which indicated the efficiency of phage therapy against Shigella strains.
-
Neonates are commonly infeced with S. marcescens, especially immunocompromised neonates of low birth weight(Larson et al., 2005). Furthermore, S. marcescens has a specific affinity for the central nervous system, and meningoencephalitis or a brain abscess with this pathogen has a severe neurologic prognosis(Messerschmidt et al., 2004). Recently, there have been reports concerning multidrug-resistant strains of S. marcescens in pediatrics, which make it difficult to treat diseases caused by this bacterium with drug therapy(Maragakis et al., 2008).
In 1967, phages were used to infect S. marcescens. Iino et al. used a broad host range phage χ to split 20 of S. marcescens strains(Iino and Mitani, 1967). Their results showed that phage χ could only infect the strains with flagella, which indicated the receptor site of phages. Matsushita et al. isolated two phages, KSP90 and KSP100, from environmental water that were related to the T4-type phage and phiEco32 phage, respectively(Matsushita et al., 2009). They extensively studied the biological features, DNA features, virion proteins and phylogenic relationships of the two phages, and their work indicated the therapeutic potential of the phages to control S. marcescens infection. Denyes et al. sequenced the genome of Serratia bacteriophage η, and have a well known about the genome length and structure, as well as all the functional CDSs(Denyes et al., 2014). Whether it carries virulence genes or not is also clear. This kind of work is useful for the application of phages for therapy.
-
Besides the above mentioned Enterobacteriaceae, studies of phages related to other Enterobacteriaceae such as Edwardsiella(Yasuike et al., 2013), Proteus(Lazareva et al., 2001), Erwinia(Born et al., 2011), and Citrobacter(Chaudhry, 2014)have also been reported recently. These works indicate the practicability of therapeutic candidates of bacteriophages.
Escherichia coli
Salmonella enterica
Klebsiella pneumoniae
Shigella strains
Serratia marcescens
Other strains of Enterobacteriaceae family
-
Bacteriophage therapy is an effective way to control bacterial infections; it is superior to antibiotic treatment in the following aspects. The phages are capable of increasing in numbers specifically where hosts are located during the bacterial-killing process, and contribute to establishing the phage dose; in addition, the cost of agent production is relatively low(Abedon and ThomasAbedon, 2010). Most phages have a specific host range, which makes them split the target pathogenic bacteria while leaving minimal disruption of normal flora(Gupta and Prasad, 2011). By contrast, many chemical antibiotics have broader spectrums of activity, which may be prone to inducing super infections(Carlton, 1999). Unlike antibiotics, which can be toxic, phages display little or no toxicity to the flora and environment(Bentley and Bennett, 2003).
Bacteriophage therapy also has limitations. One is the safety problem. All of the phages in a given cocktail will need to be appropriately characterized before they can be used in clinical treatment. Fortunately, the rapid improvement of genome sequencing technologies assures the safety of phages used in therapy approaches. Another problem is the narrow host range; however, this limitation can be counteracted by the use of recently developed and improved phage cocktails. The third limitation is the instability of the phage therapeutic agent; further study is needed to improve and prefect this factor. The forth problem is the phase-resistance developed by bacteria during their co-evolution with phages. Disputes focus on whether the same result will appear in the future, as bacteria will develop multi-phage resistance as the same as the abuse of antibiotics. This is an important problem and needs future studies.
-
This work was supported by the National Infrastructure of Microbial Resources(NIMR2014-4).
-
All the authors declare that they have no competing interests. This article does not contain any studies with human or animals subjects performed by any of the authors.

 DownLoad:
DownLoad: