









HTML
-
CTXΦ is a single-stranded filamentous DNA phage that can be horizontally transferred among Vibrio cholerae strains (Ochman et al., 2000; Waldor and Mekalanos, 1996). The gene ctxAB, carried by CTXΦ, encodes the cholera toxin (CT), which is the main causative factor of cholera. CTXΦ can be integrated into the chromosome of V. cholerae through lysogenesis. The genome size and the overall arrangement of CTXΦ genes in V. cholerae are very similar to filamentous bacteriophages f1, fd, and M13 in Escherichia coli that have different F pili specificities (Russel 1995; Waldor and Mekalanos, 1996). The typical genome size of CTXΦ is approximately 7 kb and consists of two parts: the RS2 sequence (4.6 kb in size) and the core region (2.4 kb in size; Figure 1). Three genes (rstR, rstA, and rstB) and two spacers (ig-1 and ig-2) are present in the RS2 region. RstA, encoded by rstA, is related to CTXΦ replication, while rstB, encoded by rstB, is associated with the site-specific integration of CTXΦ into the chromosome of V. cholerae. In the CTXΦ core region, psh, cep, gⅢCTX, ace, zot, ctxAB, and other genes are encoded; the first four of which are associated with the assembly and structural formation of phage particles. gⅢCTX encodes pⅢ, which is related to the recognition of V. cholerae surface receptors. Proteins encoded by zot and ace genes function in the formation of phage particles. ctxAB genes are not associated with phage formation; instead they encode toxin subunits A and B, which form the A1B5 type CT protein complex. After ctxAB -carrying CTXΦ infects V. cholerae cells, it integrates its DNA into the chromosome of V. cholerae at the attB (dif) integration site to facilitate the horizontal gene transfer of ctxAB among V. cholerae strains and the subsequent generation of new virulent strains.
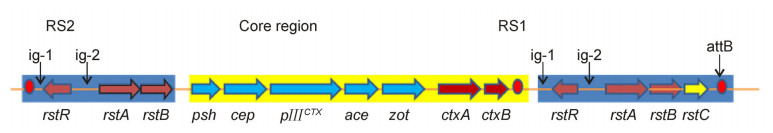
Figure 1. The genomic structure of CTXΦ in Vibrio cholerae N16961.
-
In 1999, while analyzing the genomic features of CTXΦ of El Tor type V. cholerae, we discovered that some strains did not carry the ctxAB toxin genes, though they still encoded the other genes of the CTXΦ genome. Furthermore, when drawing the physical maps of the CTXΦ genomes of the different strains, we discovered a phage strain that originated from El Tor type V. cholerae that carried only a classical-type rstRgene sequence without the ctxAB toxin genes. This unique genome was temporarily named nct-CTXclassΦ (Biao, 1999; Kan et al., 1999). Other studies also identified the genome structure of the lysogenic phage and named it pre-CTXΦ (Boyd et al., 2000). In this article, the non-ctxAB -containing CTXΦ phages are collectively referred to as pre-CTXΦ, which is the precursor form of CTXΦ. Pre-CTXΦ evolved into CTXΦ after acquiring the ctxAB genes.
As more strains were studied, more CTXΦ genomes with various gene sequence characteristics were identified and these formed the CTXΦ family. Based on various characteristics, including serogroup, biotype, variations in the rstR gene and ig-2 sequences, and whether the ctxAB genes were carried, these strains were classified into CTXclassΦ (Faruque et al., 2000) and pre-CTXclassΦ (Biao et al., 2002), derived from the classical strain of V. cholerae, CTXETΦ (Waldor and Mekalanos, 1996) and pre-CTXETΦ from the El Tor strain, and CTXcalcΦ (Davis et al., 1999) from the O139 strain. In addition toCTXETΦ, CTXclassΦ and CTXcalcΦ carried rstRET, rstRclass, and rstRcalc (Davis et al., 1999; Davis and Waldor, 2000c; Waldor and Mekalanos, 1996), and more gene sequences of the rstR gene, including rstR-4**, rstR-5, rstR6, rstR-232, and rstRZJ, were discovered (Li et al., 2003; Maiti et al., 2006; Mukhopadhyay et al., 2001; Wang et al., 2014). Moreover, our laboratory also discovered CTXΦ and pre-CTXΦ genomes that carried different types of rstR sequences in strains of the O1 and O139 serogroups and the non-O1/non-O139 serogroups (Li et al., 2014) and unpublished data).
-
CTXΦ is a lysogenic bacteriophage that does not kill the host bacteria; it is similar to the common infection process (Rasched and Oberer, 1986). After undergoing the processes of recognition of the surface receptor of V. cholerae, DNA injection, and chromosomal integration, CTXΦ phages exist inside V. cholerae cells in lysogenic or plasmid replication forms (RFs). They also undergo the process of production and release of new and mature phage particles, which in turn infect new hosts to complete the life cycle. These processes require not only proteins that are encoded by CTXΦ genes, but also the expression of related genes outside the CTXΦ genome in the V. cholerae chromosome.
-
TcpA, encoded by the tcpA gene of V. cholerae, is the major subunit of toxin-coregulated pilus (TCP), which not only plays important roles in the V. cholerae infection process as an adherence and colonizing factor (Herrington et al., 1988; Tacket et al., 1998), but also as the receptor of CTXФphage, as indicated by evidence from genetic studies (Waldor and Mekalanos, 1996). Strains that are sensitive to CTXФall had TCP pili. The TolQRA protein complex also plays an important role in the CTXΦ phage infection process (Heilpern and Waldor, 2000). It has been proposed that TolQRA complexes function in periplasm or intima based on Ff phage in E. coli (Webster, 1991). The TolA protein might play a role during infection as the second receptor of the CTXΦ phage. When filamentous fd phage infects E. coli, pⅢfd protein, encoded by fd, mediates the infection of fd as a lig and to recognize E. coli fimbriae. In CTXΦ, there is no protein homology to the pⅢfd sequence. However, based on the position of gⅢCTX in the CTXΦ genome and its similar sequence length to pⅢfd and through functional analysis, it has been postulated that pⅢCTX might exercise a function similar to that of pⅢfd (Heilpern and Waldor, 2003) and that it might act as a lig and to combine with TCP receptor to mediate CTXΦ infection. The process of the interaction between CTXФ and the surface receptor in bacterial cells likely requires two steps: first, pⅢCTX combines with the end of the TCP pili (Heilpern and Waldor, 2003), which subsequently leads to the contraction of the TCP so that the phage particles are closer to or even pass through the outer membrane of the bacterium (Russel et al., 1988; Sun and Webster, 1987). The TCP, as the first receptor, pulls the phage particle closer, which is particularly advantageous for phage infection, as it allows the host to effectively capture a specific phage (Riechmann and Holliger, 1997) while promoting effective binding between the phage and the second receptor TolA (Click and Webster, 1997; Riechmann and Holliger, 1997). TolQ and TolR also play important roles in this process, likely by forming a channel to transport some necessary substances through the endometrium (Webster, 1991).
Studies have identified polymorphisms in pⅢCTX (Bhattacharya et al., 2006; Boyd et al., 2000) and TcpA (Boyd and Waldor, 2002; Kumar et al., 2011; Li et al., 2003; Wang et al., 2014) protein sequences, with highly variable regions in the interacting protein domains (Heilpern and Waldor, 2003; Kirn et al., 2000). It has also been determined that a new tcpA allele exists in the toxigenic non-O1 non-O139 serogroups (O141, O8, O37) of V. cholerae and that the groups were sensitive to the filamentous phage CTXΦ (Boyd and Waldor, 2002), suggesting that the toxigenic non-O1 non-O139 serogroups may have evolved from the non-toxigenic strains through TCP with new functionality to acquire CTXΦ. Faced with the selection pressure of survival and the environment, V. cholerae evolved to form various types of TCP and became dominant strains because they carried certain specific TcpA sequences when facing the selection pressure of the environment. It has been reported that variation exists in the V. cholerae infection capabilities of CTXΦ carrying different types of TcpA sequences (Liu et al., 2005). The V. cholerae infection rate for CTXΦ was higher in vivothan in vitro (Liu et al., 2005), and the types of TCP from classical strains were different from those from the El Tor and O139 strains. It was noted that the phage's infection capabilities varied for different strains; the reason for this variance is currently unclear but is presumably related to phage immunity and the expression of TCP fimbriae. Perhaps in an in vivo environment, the non-toxigenic strains of V. cholerae are more easily converted to toxigenic strains.
-
The dissociation of lysogenic CTXΦ phage from, and its re-integration into, the host chromosome are both dependent on RecA (Kamruzzaman et al., 2014; Quinones et al., 2005). CTXΦ phage injects single-stranded DNA (ssDNA) into the host cell and uses the host polymerase to synthesize the complementary strand to form double-stranded DNA (dsDNA). This is either present in the form of circular plasmid pCTX or is integrated into the chromosome through the attP sequence in the CTXΦ phage genome and the attB sequence in the homologous region of the host chromosome, mediated by the host tyrosine recombinases XerC/XerD (Huber and Waldor, 2002). The phage then exists in its lysogenic form (Huber and Waldor, 2002; Waldor and Mekalanos, 1996). For V. cholerae strains that lack the CTXΦ integration site, the phage DNA exists in the form of plasmid pCTX. Genetic evidence indicates that RstB is also required for CTXΦ integration into the chromosome, but the exact molecular mechanism remains a mystery (Waldor et al., 1997). The RstB sequence has no homology to any proteins with known function; however, it has a similar sequence as LOOP, which binds to DNA and also exists in ssDNA-binding proteins (SSBs) encoded by some phage genome sequences. Some studies have found that SSB is beneficial to ssDNA stability before the ssDNA phage is packaged to become the mature phage particle (Russel, 1995).
The dissociation of lysogenic phage CTXΦ DNA from the chromosome has not yet been observed under natural environmental or growth conditions. However, it was recently reported that the CTXΦ genome sequence is adjacent to the RS1 sequence in some toxigenic El Tor strains of the O1 and O139 groups of V. cholerae. RS1 is a satellite phage related to CTXΦ, whose spread and proliferation require relevant proteins encoded by the CTXΦ genome, meaning that CTXΦ is the helper phage of RS1. Compared to RS2, the RS1 sequence only encodes one extra rstC gene (Davis et al., 2002); all the remaining genes are identical. The RS1 sequence is generally packaged together with the CTXΦ DNA sequence into phage particles (Davis et al., 2002). When strains were superinfected with RS1 phage and incubated inside small intestine ligation segments in adult rabbits, the RS1 phage caused an unstable arrangement of lysogenic CTXΦ -RS1 on the chromosome, and lysogenic CTXΦ (in some cases, together with TLC or RS1) was dissociated from the chromosome, resulting in a new non-toxigenic V. cholera; a process that was also RecA-dependent. Over-expression of the RstC protein alone in toxigenic El Tor strains of the O1 and O139 groups was sufficient to cause a similar phenomenon (Kamruzzaman et al., 2014), which led to the discovery of a new function of the RS1 phage. The RS1 phage was different from CTXΦ infection and did not result in the phenomenon of superinfection immunity. The newly produced non-toxigenic V. cholerae strain still contained the dif phage integration site and could be infected and re-integrated with a new CTXΦ phage (Kamruzzaman et al., 2014). The El Tor strain that caused the recent seventh cholera pandemic was infected with a CTXΦ that carried rstRclass genes (Ansaruzzaman et al., 2004; Nair et al., 2002). The emergence of these new types of strains might result from the loss of CTXΦ in the O1 El Tor strain and then the acceptance of a new type of phage as a recipient strain. Although no evidence has yet supported the idea that CTXclassΦ can be induced to dissociate from the chromosome, it has been demonstrated that, under the effect of chitin-induced transformation (Meibom et al., 2005), non-toxigenic O1 El Tor V. cholerae could take the CTXclassΦ DNA fragment and integrate it between the attB and attP sites (the intact integration sites left behind after the dissociation of CTXΦ) (Kamruzzaman et al., 2014).
-
Adjacent to the CTXΦ genome, the chromosomes of some V. choleraestrains contain the RS1 sequence. The origin of replication of the CTXΦ phage locates within the ig-1 sequence, at which RstA causes breakage to generate a single-stranded nick that results in the occurrence of a 3' end of DNA (Moyer et al., 2001; Waldor et al., 1997), which the host DNA polymerase uses as a template to initiate DNA synthesis (Moyer et al., 2001). When reaching the next nick at the origin of replication, the DNA synthesis stops, and the synthesis of the ssDNA that is used in the packaging of new CTXΦ phage particles completes (Moyer et al., 2001). After the newly synthesized CTXΦ phage DNA is packaged into the protein capsid, the mature phage particle progeny are assembled. Classical O1 V. cholerae cannot produce infectious CTXΦ phage particles, likely due to one of two possibilities: either the CTXΦ phage genome is present alone on the chromosomes of these strains (i.e., RS1 sequences are absent from the adjacent sequences) or two incomplete phage genomes reside on the chromosome in tandem (Davis and Waldor, 2000c), which indicates that the arrangement of CTXΦ -RS1 is critical for the generation of mature progeny phage.
-
Similar to other filamentous phages, the extracellular release of CTXΦ does not cause host cell lysis. PI, the membrane protein of Ff phage, plays an important role in the assembly and secretion of Ff phage; the Zot protein of CTXΦ phage is homologous to PI (Koonin, 1992; Waldor and Mekalanos, 1996) and contains an ATPase domain, which perhaps provides energy for the assembly and release of CTXΦ phage. CTXΦ is secreted through the channel made by the outer membrane protein EpsD, encoded by the host's T2SS secretion system (Davis et al., 2000a). EpsD is an important constituent of T2SS in V. cholerae. T2SS consists of 15 protein types and is related to the secretion of toxin CT, hemagglutinin-protease, chitinase, and other proteins (Connell et al., 1998; Sandkvist, 2001; Sandkvist et al., 1997). However, CTXΦ phage release only involves EpsD (Davis et al., 2000a).
After the integration of CTXΦ onto the V. cholerae chromosome, the phage makes use of proteins encoded by the host genome and undergoes replication, assembly, and secretion to complete its life cycle, but it does not cause the lysis and death of V. cholerae cells. It also does not affect the growth of the bacterium; in contrast, the CTXΦ and V. cholerae co-evolve. CTXΦ carries the CT gene, and CT causes diarrhea in infected patients to promote the propagation and proliferation of V. cholerae, indicating that the relationship between CTXΦ and its host V. cholerae is mutually beneficial.
-
The promoter (PrstA) of the rstA gene inside the RS2 region regulates the overall transcription of genes that are associated with CTXΦ phage replication and morphogenesis. The direction of rstR transcription is opposite to that of rstA, and there is a spacer sequence (ig-2) between rstR and rstA, which contains PrstA and the rstR operon. The RstR protein binds to the ig-2-binding region upstream of the rstA open reading frame (ORF) to inhibit rstA transcription to maintain CTXΦ lysogenesis (Kimsey and Waldor, 1998; Waldor et al., 1997). In addition to phage-encoded RstR, the host's SOS reaction regulatory protein LexA can also bind to PrstA to inhibit the transcription of downstream genes (Kimsey and Waldor, 2009; Quinones et al., 2005). Under normal culture conditions, both LexA and RstR bind to PrstA to inhibit its transcription (Quinones et al., 2005). Acting in a tetramer, RstR binds to three different sites O1, O2, and O3 (each site approximately 50 bp in size) of the RstA gene promoter, with the tightest binding at the O1 site and relatively weak binding at the O2 and O3 sites. The O2 binding site overlaps the −10 to −35 nt positions of the RstR promoter (PrstR), suggesting that RstR may inhibit its own transcription (Kimsey and Waldor, 2004). The SOS reaction conditions that cause DNA damage will increase the induction of CTXΦ phage particles (Quinones et al., 2005). The SOS reaction caused by mitomycin C and ultraviolet (UV) light leads to increased activity of the auxiliary protease RecA, which is related to the DNA repair pathway, which in turn causes the self-degradation of LexA, the global regulatory factor of the SOS reaction. After LexA degradation, the RstR protein level is reduced, which lifts the transcriptional repression on RstA (Quinones et al., 2005) and ultimately leads to increased production of CTXΦ phage (Quinones et al., 2005; Waldor and Mekalanos, 1996). Studies have shown that after the mitomycin C-induced inhibition on PrstA was lifted, the mRNA expression of ctxA increased by seven-fold (Quinones et al., 2005). However, the increase in the CTXΦ phage particles elicited from the SOS response was limited (Quinones et al., 2005). The molecular mechanism of RstR's inhibition on RstA transcription still needs further investigation.
While RS1 utilizes proteins encoded by CTXΦ to assemble phage particles, RstC also plays a positive role in the induction of phage production. RstC directly binds to RstR to block the binding of RstR to PrstA and ultimately assists in CTXΦ proliferation (Davis et al., 2002); in other words, RS1 is a helper phage to CTXΦ. Meanwhile, PrstA controls RstC expression; thus, the factors that can enhance the RstA transcription level will also lead to enhanced RstC expression. Meanwhile, the enhanced PrstA activity will also lead to up-regulated transcription of the downstream genes of ctxAB to achieve the RstC's regulation on virulence genes (Davis et al., 2002). However, the regulation by genes from CTXΦ itself on ctxAB transcription is limited (Davis et al., 2002). Stx toxin, encoded by the stx gene in the λ phage of E. coli, is the major causative agent of enterohemorrhagic E. coli (EHEC), in which the production and release of Stx toxin is mainly dependent on inhibitors encoded by the λ phage itself (Neely and Friedman, 1998). Different from the regulation of the stx gene, the regulation of the ctxA gene mainly relies on the regulation of the promoter by the cell transcription factors ToxR, ToxT, and TcpPH, which are outside of CTXΦ (Krukonis and DiRita, 2003).
The RS1 sequence was present in the recently prevalent O1 El Tor and O139 strains but absent in the O1 classical strain that caused the sixth cholera pandemic (Davis et al., 2000b; Waldor et al., 1997). It is possible that the acquisition of the rstC gene conferred an advantage to the recently prevalent strains (Dziejman et al., 2002). In summary, there exists a fascinating and mutually supplementary relationship between the satellite phage RS1 and CTXΦ; proteins encoded by CTXΦ are required for the formation of phage particles for RS1, while RS1 is beneficial to CTXΦ during both its dissociation from the chromosome and proliferation.
The integration and dissociation of CTXΦ phage on the host chromosome
The replication of CTXΦ phage in host cells
The assembly and release of CTXΦ phage
The regulatory mechanisms of CTXΦ phage induction
-
V. choleraehas two chromosomes, which are 2.9 and 1.1 Mb in size. While CTXΦ can be integrated into the large chromosome, it can also be integrated into the small chromosome in the classical strain of the O1 group (Davis et al., 2000b) and in some El Tor strains (Nandi et al., 2003) prior to the emergence of the O139 strains. For the El Tor N16961 strain of the O1 group of V. cholerae, which was the first strain for which whole-genome sequencing was completed (Heidelberg et al., 2000), its CTXΦ resides on the large chromosome, while on the small chromosome there is a single empty integration site that is similar to the one on the large chromosome. In classical strains of the O1 group, CTXΦ can be present on both the large and the small chromosome (Davis et al., 2000b; Trucksis et al., 1998). Additionally, an integrated CTXΦ genome exists on the small chromosomes of some E1 Tor strains (Nandi et al., 2003). The regions of variation in different types of CTXΦ phages in the CTXΦ family are mainly concentrated in the rstR-ig2 sequences in the RS region; therefore, the rstR-ig2 sequence serves as the main basis for distinguishing different alleles in the phage family. Different alleles of the CTXΦ family can be integrated into the same strain.
Homologous recombination events exert great influence on the arrangement of CTXΦ on the host chromosome. During the CTXΦ integration process, homologous recombination can occur, not only in pCTXΦ which exists as RF, and between the attB integration sites on the chromosome, but also in between the important elements related to the CTXΦ genome. It was recently determined that the occurrence of an atypical El Tor strain of V. cholerae (with biochemical characteristics of the El Tor strain of V. cholerae, except that the CT gene sequence was not the one that is typically carried by the El Tor strain) may have originated through interchromosomal or intrachromosomal homologous recombination in the relevant homologous regions from an intermediate strain that was infected by different types of CTXΦ phages (Kim et al., 2014). This recombination did not involve large changes to the chromosome, but rather limited changes to small relevant elements related to the CTXΦ phage, such that the biochemical characteristics of the strain remained unchanged. The outcome of the recombination altered the arrangement of CTXΦ on the large and small chromosomes, representing an important event in V. cholerae evolution. Such events may also be one of the important reasons for the occurrence of chimera phages.
The presence of CTXΦ in V. cholerae is complex, with varying CTXΦ types, copy numbers, and polymorphisms of position and arrangement on the two host chromosomes, which are related to the RS region that encodes the dissociation and integration functions. It is affected by factors such as the attB site and its adjacent sequences, such as TLC. All of these factors are related to the transfer and integration of CTXΦ in V. cholerae and, therefore, to the evolution of V. cholerae pathogenicity. The evolution of the strain is a very long process, and the long-lasting interactions among many factors lead to polymorphisms. The mechanism of interactions between CTXΦ and other factors and the evolutionary direction of strains requires further in-depth study.
-
This study was supported by the State Key Laboratory for Infectious Disease Prevention and Control of China (Grant number 2014SKLID101) and the Priority Project on Infectious Disease Control and Prevention (2012ZX10004215).
-
All the authors declare that they have no competing interests. This article does not contain any studies on human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: