












HTML
-
Baculoviridae is a family of rod-shaped, enveloped, insect-specific viruses with a large, covalently closed, double-str and ed DNA molecule ranging in size from 80 to 180 kilobases(Rohrmann, 2013). Based on the sequenced genomes and phylogenetic relationships within the family, Baculoviridae is subdivided into four genera: Alphabaculovirus, Betabaculovirus, Gammabaculovirus and Deltabaculovirus(Jehle et al., 2006). Betabaculovirus, which encompasses only lepidopteran-specific granuloviruses(GVs), is separated into three groups according to the pathological features of the insect hosts. Type Ⅰ GV pathology is characterized by an infection constrained to the host's midgut and fat body and a relatively slow killing speed. Type Ⅱ GVs are able to infect most of the host's tissues and exhibit a rapid killing. The third pathology, with a single representative, Harrisina brillians granulovirus, causes an infection confined to the midgut epithelium, resulting in the rapid death of the host(Federici, 1997). Because permissive cell lines are limited, GVs have been less well studied than lepidopteran-specific nucleopolyhedroviruses(NPVs), which belong to Alphabaculovirus (Winstanley and Crook, 1993).
Cnaphalocrocis medinalis granulovirus(CnmeGV)was first isolated from Enping county, Guangdong province, China in 1979(Pang et al., 1981). The CnmeGV strain is detectable in the population of C. medinalis larvae in situ even after 30 years since its first isolation(Zhang et al., 2014). As a highly specific and effective viral pathogen, CnmeGV is one of promising biological pesticides for controlling the rice pest. Studies on CnmeGV genome will be helpful for better understanding of its pathology and utilization. Here, we present the complete sequence and organization of the CnmeGV genome, and compare them to other baculoviruses using genomic and phylogenetic analyses.
-
The strain of CnmeGV, designated as Enping strain, was isolated in 1979 and stored in our lab(Pang et al., 1981). The occlusion bodies(OBs)of CnmeGV were propagated in C. medinalis larvae and then were purified using differential centrifugation(O'Reilly et al., 1992). To extract viral DNA, the purified OBs were resuspended in 0.1 mol/L sodium carbonate solution [0.1 mol/L Na2CO3, 0.17 mol/L NaCl, 0.01 mol/L EDTA(pH 10.9)] and incubated at 37℃ overnight with 0.5 mg/mL proteinase K(Sigma-Aldrich®Shanghai, China) and 1% SDS. After extraction with phenol:chloroform: isoamyl alcohol(25:24:1, v/v), the viral DNA was precipitated with ethanol and then resuspended in TE buffer(10 mol/L Tris-HCl, pH 8; 1 mol/L EDTA).
-
The genome was sequenced with the Roche 454 GS FLX system by using shotgun strategy. The reads were assembled with Roche GS De Novo assembler software. Contigs assembly was performed with assistance of endonuclease restriction analyses. The genome sequence data was submitted to GenBank(http://www.ncbi.nlm.nih.gov/genbank).
Hypothetical ORFs were predicted by soft berry FGENESV program(http://www.softberry.com/berry.phtml)(Solovyev and Salamov, 1999) and defined by the standard ATG start, and a stop codon and potentially encode at least 50 amino acids. Gene annotation and comparisons were done with NCBI protein-protein BLAST algorithm(http://blast.ncbi.nlm.nih.gov/Blast.cgi). Repeat structures were detected by BLAST alignment of two sequences(http://blast.ncbi.nlm.nih.gov/Blast.cgi). The identity among homologous genes was calculated with MegAlign software using ClustalW with default parameters. Restriction sites were predicted by MapDraw software. Genome map framework was drawn with genomeVX(Conant and Wolfe, 2008).
-
The phylogenetic analysis was performed based on amino acid sequences of 37 core genes form CnmeGV and the other 63 baculoviruses listed in the NCBI genome database(http://www.ncbi.nlm.nih.gov/nuccore/?term=baculovirus). All the sequences were linked by a stationary order and multiple alignments using ClusterW method with MEGA5 by using default settings. A phylogenetic tree was constructed by MEGA5 using Maximum Likelihood method based on the JTT matrix-based model(Jones et al., 1992; Tamura et al., 2011). Phylogeny was tested by Bootstrap method with a value of 1000(Sanderson and Wojciechowski, 2000).
Viral DNA extraction
Sequencing and bioinformatic analysis
Phylogenetic analysis
-
To date, 74 baculovirus genomes have been fully sequenced(http://www.ncbi.nlm.nih.gov/genomes/GenomesGroup.cgi?taxid=10442)(Liu et al., 2014; Yin et al., 2015) and 18 of them belong to the Betabaculovirus genus. In the present study, the genome of CnmeGV was sequenced using the Roche 454 GS FLX system with shotgun strategy. Totally 692.5 times coverage of the genome was achieved by using the generated 77, 041, 934 nucleotides(nt)of raw data from 150, 856 sequencing reads. The size of the assembled CnmeGV genome is 111, 246 nt. It has a high A+T content of 64.8% which is similar to that of Phthorimaea operculella granulovirus(64.3%)(PhopGV, GenBank accession no. NC_004062) and Adoxophyes orana granulovirus(65.5%)(AdorGV, GenBank accession no.NC_005038)(Taha et al., 2000; Wormleaton et al., 2003). The coding sequences coverage 93% of the CnmeGV genome.
In total, 118 putative open reading frames(ORFs)were identified with at least 50 codons in length and minimal overlap. The granulin gene was defined as the first ORF and the adenine of its initiation codon was defined as the first nucleotide of the genome. As compared with other baculovirus genomes, the ORFs randomly distributed with 63 in the granulin-sense orientation and 55 in the opposite orientation. Among these ORFs, it contains the 37 conserved baculovirus core genes, 13 unique genes, 26 genes found in all sequenced lepidopteran baculoviruses, and 42 common genes that are present in a variety of baculoviruses(Figure 1).
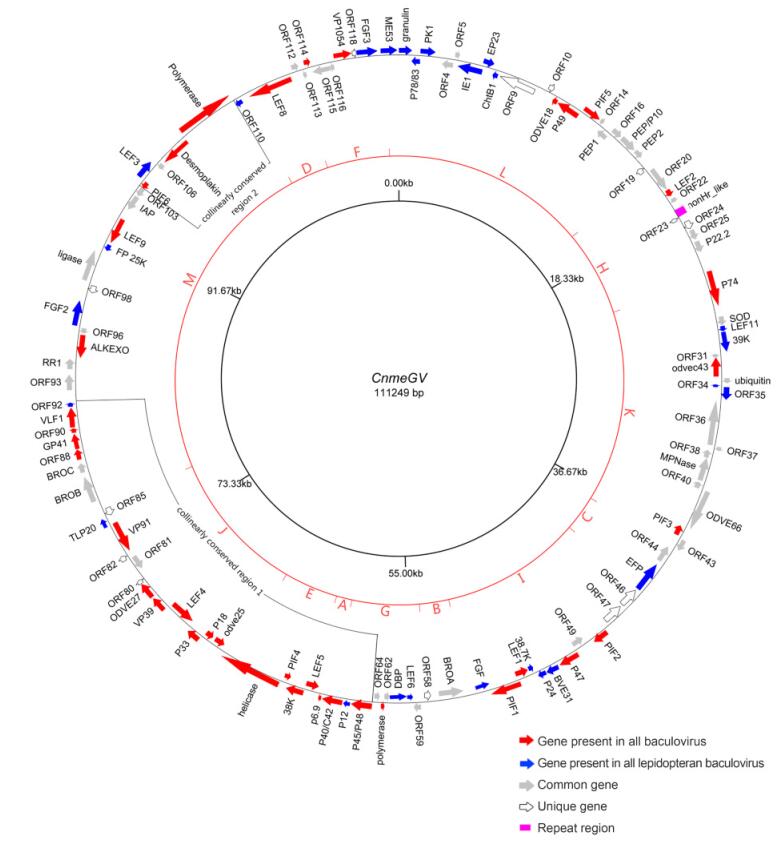
Figure 1. Genome map of CnmeGV. ORFs are indicated by arrows with a displayed name. Arrows also signify transcription directions. Red arrows represent core genes, blue represent genes present in all lepidopteron baculoviruses, gray represent baculoviral common genes and open arrows represent unique genes of CnmeGV. The pink square represent a repeat structure. The inner circle indicates genome scale position by 20 kb. Hind Ⅲ restriction map is shown in the middle dark red circle. A region collinearly conserved in alpha-and betabaculoviruses is also shown.
-
Phylogenetic analysis was performed based on 37 concatenated core genes of 64 representing baculovirus genomes(Figure 2). The obtained clad gram shows four groups of different genera, which were recognized in the current classification of the family(Jehle et al., 2006). CnmeGV is grouped in Betabaculovirus genus as expected. It forms a big clade with other seven GVs including AdorGV, Clostera anastomosis granulovirus(CaLGV), Choristoneura occidentalis granulovirus(ChocGV), Clostera anachoreta granulovirus(ClanGV), Cydia pomonella granulovirus(CpGV), Cryptophlebia leucotreta granulovirus(CrleGV) and Pieris rapae granulovirus(PrGV).
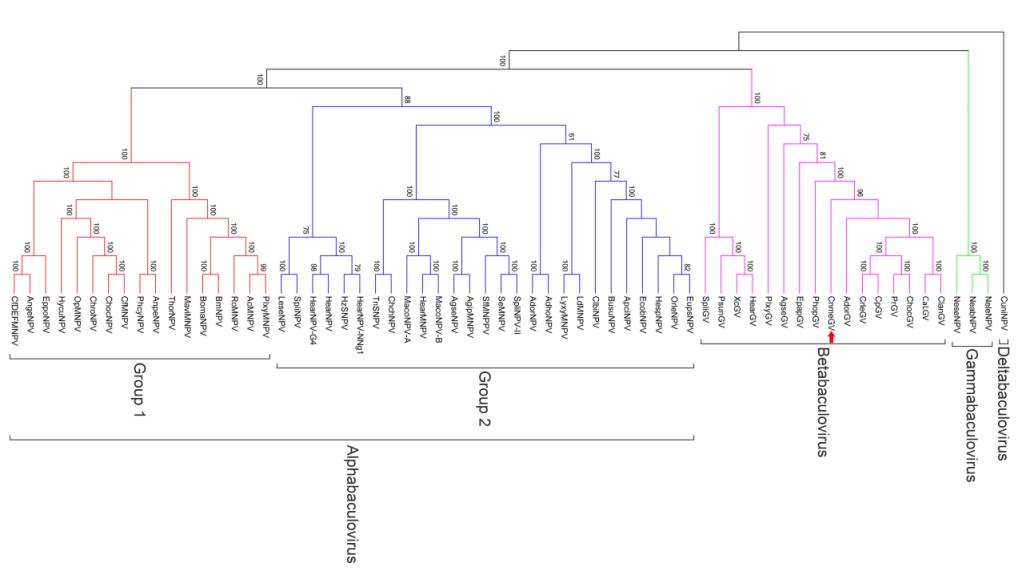
Figure 2. Phylogenetic tree using 37 core proteins of 64 sequenced baculoviruses based on Maximum Likelihood method. It tested by Bootstrap method with a value of 1000. The bootstrap values greater than 50% are showed in front of every nodes. Arrow points to CnmeGV.
By using Gene Parity Plot(Hu et al., 1998), the gene colinearity of CnmeGV was compared to other sequenced GVs. The result showed that CnmeGV differs from the rest of the GVs by a 23 kb gene block inversion from Cnme25 to Cnme48 and a 17kb gene block inversion from Cnme92 to Cnme108. Gene-parity plots of CnmeGV with four closely related GVs including AdorGV, ClanGV, CpGV and PrGV based on ORF order are shown in Figure 3.
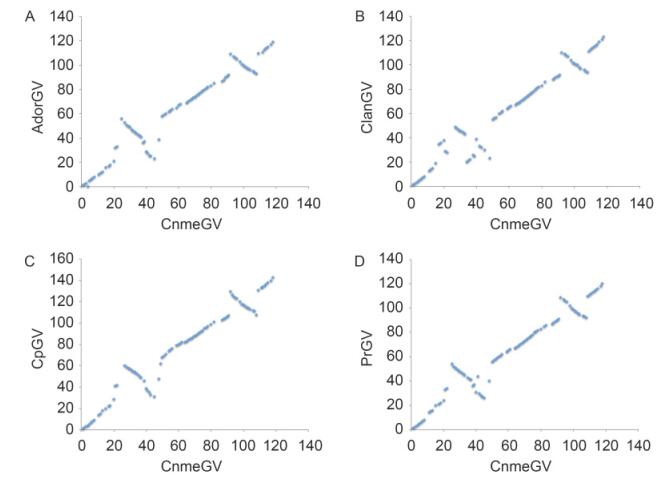
Figure 3. Gene-parity plots of CnmeGV with AdorGV (A), ClanGV (B), CpGV (C) and PrGV (D) based on ORF order.
The nucleotide identities between the ORFs of CnmeGV and other representative sequenced GVs are shown in Supplementary Table S1. There are 72–95 homologous ORFs between CnmeGV and the 15 typical sequenced GVs, and the maximum and minimum homologies are with PrGV(95) and Plutella xylostella granulovirus(PlxyGV)(72), respectively. The genes present or missing in CnmeGV genome are summarized in Table 1.
Gene function Names of genes presented in CnmeGV(ORF numbers of CnmeGV) Genes missing in CnmeGV Transcription (9) lef-11(29), 39k(30), p47(50), lef-6(60), lef-5(69), lef-4(76), vlf-1(90), lef-9(100), lef-8(109) pe-38 Replication (12) ie-1(6), lef-2(21), lef-1(54), dbp(61), dnapol(63, 107), helicase(72), rr1(93), dnaligase(98), lef-3(104), me53(118) rr2 Structure (30) granulin(1), p78/83(2), pk-1(3), odv-e18(11), pep-1(15), pep/p10(17), pep-2(18), odv-ec43(32), odv-e66(41), efp(45), BV-e31(51), p24(52), p6.9(68), odv-e25(73), p18(74), p33(75), vp39(77), odv-e27(78), tlp20(83), gp41(88), fp25k(99), vp1054(115) -- Per os infectivity pif-5(13), p74(27), pif-3(42), pif-2(48), pif-1(55), pif-4(71), vp91 (82), pif-6(103) -- Auxiliary (11) sod(28), ubiquitin(33), mp-nase(39), fgf-1(56), bro-A(57), bro-B(85), bro-C(86), alk-exo(94), fgf-2(96), iap-5(101), fgf-3 (117) chitinase, cathepsin, ctl, p35, lef-10, egt, gp37, enhancin Unknown (45) 4, 5, 7, 8, p49(12), 14, 16, 20, 22, 25, p22.2 (26), 31, 34, 35, 36, 37, 38, 40, 43, 44, 49, 38.7k (53), 59, 62, 64, p45/48(65), p12(66), p40/c42(67), 38k(70), 80, 87, 89, 91, 92, 95, 102, 105, desmoplakin (106), 108, 110, 111, 112, 113, 114 -- CnmeGV unique (13) 9, 10, 19, 23, 24, 46, 47, 58, 79, 81, 84, 97, 116 -- Note: The ORFs that are presented in all sequenced baculoviruses are printed in bold. Table 1. CnmeGV genes grouped according to function.
-
Homologous repeated sequences(hrs)were supposed to be characteristic for many baculovirus genomes. Hrs seem absent from the CnmeGV genome, but a repeat sequence, which is probably able to form a hairpin structure, was detected in the CnmeGV genome from nt 18102 to 18575 with a high A/T content(75.5%)(Supplementary Table S1).
-
Several genes required for baculovirus late gene transcription have been described(Lu and Miller, 1997). Like all other baculoviruses, CnmeGV codes for all four subunits of the viral encoded RNA polymerase, lef-4 (Cnme76), lef-8 (Cnme108), lef-9 (Cnme100) and p47 (Cnme50). Two other core genes, lef-5 (Cnme69) and vlf-1 (Cnme90), and three non-core genes, lef-11 (Cnme29), 39k (Cnme30) and lef-6 (Cnme60), which may be also related to the transcription process, were found in the CnmeGV genome(Table 1). Of the baculoviral early transcription genes ie-0, ie-1, ie-2 and pe38, only ie-1 (Cnme6) is present in the CnmeGV genome and is poorly conserved(about 35.7% averaged amino acid identity to those of other GVs).
-
Six viral genes(lef-1, lef-2, lef-3, dnapol, helicase and ie-1)have been identified as essential genes for baculovirus DNA replication(Lu et al., 1997; Lu and Miller, 1997). Homologues of all these genes were found in the CnmeGV genome. It is noteworthy that CnmeGV has two dnapol genes(Cnme63 and Cnme107). Cnme63 encodes 62 amino acids and has only low identity to several NPVs whereas Cnme107 codes for 1086 amino acids and has homologues to all sequenced GVs with high identity.
Genes for enzymatic functions in nucleotide metabolism such as the large(rr1) and the small(rr2)subunits of ribonucleotide reductase(RNR) and deoxyuridyltriphosphate(dUTPase)have been reported in baculovirus genomes(Taha et al., 2000; Luque et al., 2001; Ferrelli et al., 2012; Zhang et al., 2014). These enzymes can catalyze the reduction of host cell ribonucleotide diphosphates to yield deoxyribonucleotides(Lange and Jehle, 2003). Of these genes, CnmeGV contains only the rr1 (Cnme93) gene, which has been also found in Agrotis segetum granulovirus(AgseGV), CpGV, Epinotia aporema granulovirus(EpapGV) and PhopGV(Taha et al., 2000; Luque et al., 2001; Ferrelli et al., 2012; Zhang et al., 2014).
CnmeGV also encodes a DNA ligase(dnaligase, Cnme98) which is present in all sequenced GVs. dnaligase seems to be linked to the appearance of a second helicase, helicase-2 (Herniou et al., 2003), which is not found in CnmeGV. Other genes related to DNA replication, including DNA binding protein(dbp, Cnme61) and me-53(Cnme118), have also been identified in the CnmeGV genome.
-
In the CnmeGV genome, 19 core structural genes were identified, such as the core protein, p6.9(Cnme68); the tegument protein, gp41(Cnme88); the capsid-associated proteins, 38k(Cnme70), vp39 (Cnme77) and vp1054(Cnme115); the occlusion-derived virus(ODV)envelope proteins, odv-e18 (Cnme11), odv-e25 (Cnme73) and odv-e27(Cnme78); the per os infectivity factors p74(Cnme27), pif-1 (Cnme55), pif-2 (Cnme48), pif-3 (Cnme42), pif-4(Cnme71), pif-5(Cnme13), pif-6(Cnme103) and vp91(Cnme82). Other core genes, including p18 (Cnme74) which are related to the viral nucleocapsid and p33 (Cnme75) which encodes a type of a sulfhydryl oxidase(Wu and Passarelli, 2010), were also identified.
Some non-core structural genes were found in CnmeGV genome, including granulin (Cnme1), p78/83 (Cnme2), pk-1 (Cnme3), pep-1 (Cnme15), pep/p10 (Cnme17), pep-2 (Cnme18), odv-e66 (Cnme41), efp (Cnme45), BV-e31 (Cnme51), p24 (Cnme52), tlp20 (Cnme83) and fp25k (Cnme99), but CnmeGV lacks a homologue of odv-e56.
-
Auxiliary genes are not essential for viral replication, but they can provide a selective advantage(O'Reilly, 1997). The CnmeGV genome contains neither a chitinase nor cathepsin gene(Table 1). That may explain why CnmeGV infected insects do not show the phenotype of typical liquefaction. Among the auxiliary genes, ubiquitin is the most conserved one and it has been found in all sequenced GVs. Cnme33 is an ubiquitin homologue, showing 80% mean amino acid identity to those of the sequenced GVs(Supplementary Table S1). All of the sequenced GV genomes encode three fibroblast growth factors(fgfs), and CnmeGV is not an exception, with Cnme56, 98 and 117 at the corresponding locations as seen in other GVs. CnmeGV also contains a sod(Cnme28) and a iap gene(iap-5, Cnme101). Three baculovirus repeated orf(bro)genes have also been found in CnmeGV including bro-a(Cnme57), bro-b(Cnme85) and bro-c(Cnme86). The absence or duplication of these genes is common in baculovirus(Zhou et al., 2012). BRO has been demonstrated to possess DNA binding activity, especially to single str and ed DNA(Zemskov et al., 2000).
Sequencing and general characteristics of the CnmeGV genome
Relationships with other baculoviruses
CnmeGV genes grouped according to function
Repeat structure
Transcription genes
Replication genes
Structural genes
Auxiliary genes
-
Rice is a very important crop all over the world. The Asiatic rice leafroller, Cnaphalocrocis medinalis(Lepidoptera: Pyralidae), is the main migratory pest of rice with 1–11 generations per year, depending on its geographical distribution in rice-planting regions(Chai et al., 2011). The vast use of chemical insecticides for controlling C. medinalis results in increased insect resistance and environment pollution. After first isolated in C. medinalis larvae in 1979, CnmeGV still existed stably in the C. medinalis population in situ for more than 30 years(Zhang et al., 2014), which suggests that CnmeGV is a potential specific and effective pathogen to control the target pest in field. The genomic analyses of CnmeGV will be of great helpful for developing it as a biological pesticide for the control of C. medinalis.
Strong colinearity is observed in GV genomes sequenced to date(Luque et al., 2001; Lange and Jehle, 2003; Wormleaton et al., 2003; Hilton and Winstanley, 2008). However, CnmeGV differs from the rest of sequenced GVs by a 23 kb and a 17 kb gene block inversions. The first inversion begins with Cnme25 and ends with Cnme45. Except 3 core genes(p74, odv-ec43, pif-3), most of genes in this region are baculovirus common genes. The secondary inversion contain genes from Cnme92 to Cnme108. Four of the baculovirus core genes can be found in this region: alk-exo, lef-9, pif-6 and desmoplakin. Baculovirus genes with different functions are usually scattered throughout the genomes and ORFs are present on both strands of the DNA, so does the genes within the two fragments. Although the gene block inversion is uncommon in GVs, it is a common feature in NPVs, even in a same phylogenetic clade. No research result shows the function or effect of the inversion.
Homologous repeated sequences(hrs)are found in most baculovirus genomes with varying length(Ferrelli et al., 2012). Hrs were suggested to be origins of viral DNA replication(Kool et al., 1993; Hilton and Winstanley, 2008) and enhancers for transcription(Guarino et al., 1986; Choi and Guarino, 1995; Rodems and Friesen, 1995). However, the CnmeGV genome does not contain typical baculovirus hrs but probably contain a hairpin structure. A non-hr origin that contains palindromic and repetitive sequences in a complex organization was also demonstrated to initiate replication(Kool et al., 1993; Wu and Carstens, 1996). Whether this is a functional non-hr origin for CnmeGV needs further analysis.
Auxiliary genes are also compared and analyzed in this study. Chitinase and cathepsin genes are related to the breakdown of insect tissues at the end of infection. Deleting either the chitinase or cathepsin gene of AcMNPV results in the failure of host liquefaction(Slack et al., 1995; Hawtin et al., 1997). The absent of chitinase and cathepsin genes in CnmeGV genome explains why infection of CnmeGV cannot result in liquefaction of the host insects in a typical manner(Zhang et al., 2014).
In summary, the complete genome of CnmeGV was sequenced and analyzed in the present study. These results suggested that CnmeGV is a distinct species in Betabaculovirus and has closer relationship to AdorGV than to the other baculoviruses. CnmeGV genome differs from the rest of sequenced GVs by a 23 kb and a 17 kb gene block inversions. It does not contain typical baculovirus hr, but probably contains a new repeat structure. More interestingly, chitinase and cathepsin genes, which are reported to have major roles in the liquefaction of the hosts, were not found in the CnmeGV genome. This study, along with future studies about the characterization of CnmeGV infection, will be useful for the better understanding of the pathology caused by this virus and its potential utilization as a bioinsecticide.
-
This research was supported by the Hi-Tech Research and Development Program of China(863 Program; grant 2011AA10A204) and the State Key Laboratory of Biocontrol(SKLBC13KF01). The authors also thank the 454 sequence services from the core facility center of Wuhan Institute of Virology, CAS.
-
The authors declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by any of the authors.
-
KY and FD designed the experiments. SZ, ZZ, SS and QC carried out the experiments. SZ, ZZ, FD, and KY analyzed the data. SZ, FD and KY wrote the paper. All authors have read and approved the final manuscript.
Supplementary Table S1 is available on the website of Virologica Sinica: www.virosin.org; link.springer.com/journal/12250.
-
CnmeGV Homologous ORFs in GVs (ORF number/amino acid identity based on Blastp results %) orf start end aa name Ador Agse CaL Choc Clan Cp Crle Epap Hear Phop Pira Plxy Psun Splt Xecn 1 1 > 750 249 granulin 1/83 1/80 1/83 1/83 1/83 1/84 1/84 1/84 1/80 1/82 1/76 4/79 1/79 1/79 1/80 2 747 < 1220 157 p78/83 2/33 2/52 2/39 2/43 2/41 2/41 2/35 5/33 2/29 2/51 2/29 3 1207 > 2016 269 pk-1 3/52 3/46 3/50 3/55 3/52 3/48 3/55 6/42 3/45 3/52 3/58 6/46 3/46 3/43 3/43 4 2452 < 3081 209 4/26 6/28 4/31 5/35 4/33 4/35 4/36 4/26 6/27 4/33 4/34 8/20 6/26 5/29 7/27 5 3071 > 3346 91 5/38 7/31 5/43 6/42 5/48 5/42 5/40 5/32 5/43 9/30 6/39 6 3326 < 4720 464 ie-1 6/43 8/32 6/39 7/43 6/38 7/40 6/37 35/32 8/31 6/36 6/46 10/29 8/30 7/28 9/31 7 4654 > 5238 194 ep23 7/41 9/42 7/48 8/44 7/47 8/42 7/49 34/41 9/31 7/38 7/54 11/32 9/33 8/31 10/31 8 5339 < 5662 107 chtB1 8/47 10/40 8/50 9/51 8/50 9/55 8/55 33/47 10/42 8/49 8/52 12/53 10/42 9/46 11/42 9 5820 < 7844 674 U 10 8455 > 8739 94 U 11 8980 < 9261 93 odv-e18 10/74 11/52 13/74 12/86 13/73 14/77 13/78 29/73 11/58 12/76 14/81 13/69 11/57 10/63 12/60 12 9262 < 10650 462 p49 11/49 12/46 14/51 13/56 14/51 15/57 14/57 28/43 12/40 13/38 15/60 14/41 12/39 11/41 13/40 13 10678 > 11775 365 pif-5 12/66 15/57 15/71 14/74 15/67 18/70 17/69 27/68 14/51 16/68 16/67 16/64 14/53 13/53 15/50 14 11776 > 11994 72 19/40 15 11989 < 12624 211 pep-1 16/44 18/48 19/53 17/59 19/54 20/50 20/55 25/51 16/50 19/52 20/51 20/66 16/51 15/46 17/50 16 12614 > 13186 190 9/38 33/32 10/32 8/36 10/31 11/31 17 13278 > 14228 316 pep/p10 17/53 19/49 36/52 18/57 35/53 22/51 23/55 22/52 18/59 20/56 21/56 21/47 18/60 19/40 19/59 18 14264 > 14728 154 pep-2 18/40 20/47 37/54 19/49 36/53 23/48 24/49 21/42 17/46 21/50 22/48 23/46 17/46 18/42 18/47 19 15143 < 15415 90 U 20 15606 > 16982 458 21/29 40/25 38/25 29/30 24/24 21 16985 > 17392 135 lef-2 32/27 35/27 30/40 29/33 29/28 41/45 38/31 33/36 37/46 33/53 33/39 32/26 35/27 22 17525 > 17767 80 33/42 36/41 29/45 30/46 28/49 42/40 39/40 38/37 34/45 34/38 33/38 36/37 18102 > 18575 non-hr like 23 18523 < 18693 56 U 24 19000 > 19422 140 U 25 19491 > 20090 199 56/48 57/50 48/51 59/56 54/54 26 20174 > 20809 211 p22.2 64/26 52/33 27 21892 > 23865 657 p74 53/52 56/48 52/52 46/58 49/54 60/58 58/58 59/51 72/41 55/57 51/56 49/51 77/43 77/43 28 24446 > 24946 166 sod 51/59 54/59 51/60 44/66 48/61 59/66 57/62 58/60 63/59 54/62 50/71 64/61 68/54 29 24972 > 25295 107 lef-11 50/52 52/54 50/59 43/64 47/64 58/60 56/60 57/42 51/51 53/58 49/61 46/48 54/57 49/49 56/54 30 25258 > 26397 379 39k 49/31 51/26 42/28 46/28 57/45 55/24 56/32 52/31 48/36 31 26431 < 26604 57 47/47 49/47 47/33 41/51 45/33 56/62 54/54 49/38 51/40 47/62 54/38 32 26627 < 27700 357 odv-ec43 46/54 48/57 46/55 40/60 44/54 55/52 53/57 53/53 48/41 50/46 46/66 43/48 51/41 46/48 53/41 33 27790 > 28074 94 ubiquitin 45/76 47/73 45/80 39/83 43/80 54/80 52/84 52/83 47/83 49/76 45/84 42/77 50/83 45/80 52/83 34 28130 < 28276 49 44/45 20/56 38/68 20/56 53/61 51/51 46/53 48/55 41/50 48/47 43/55 51/53 35 28291 > 28980 229 43/74 46/69 21/60 37/84 21/59 52/81 50/70 50/69 45/67 47/74 43/83 40/53 47/65 42/64 50/67 36 28990 < 31503 837 42/33 45/30 22/28 22/27 50/38 47/48 46/38 42/30 41/26 37 31531 > 31731 66 41/42 44/33 49/40 46/45 45/45 41/41 44/40 46/38 38 31798 < 32268 156 36/33 40/31 27/28 32/32 26/30 36/33 39 32253 < 33578 441 mp-nase 37/32 41/34 26/39 33/40 25/37 46/38 43/41 44/34 38/29 41/35 37/43 35/28 38/30 37/30 40/29 40 33644 < 34045 133 29/58 34/40 41/52 28/54 39/53 39/56 36/58 40/55 34/58 31/53 31/43 32/42 31/37 34/42 41 34012 > 36279 755 odv-e66 28/60 33/37 27/66 37/68 35/62 39/52 150/40 33/67 44/54 30/52 156/41 149/41 42 36280 < 36864 194 pif-3 26/38 29/36 34/46 26/42 33/45 35/45 34/47 38/42 30/43 31/42 30/47 29/41 30/46 27/46 32/44 43 36924 > 37577 217 25/24 27/29 33/21 24/29 32/24 33/24 32/24 37/26 28/32 29/29 28/30 28/28 28/27 26/27 44 37669 < 38604 311 27/37 45 38798 < 40582 594 efp 23/41 25/48 31/44 23/53 30/45 31/52 30/53 14/42 26/31 27/49 26/55 26/41 26/32 24/38 27/32 46 40704 < 41780 358 U 47 41925 < 43061 378 U 48 43398 > 44297 299 pif-2 39/64 43/50 24/67 35/69 23/67 48/70 45/67 47/61 42/55 44/66 40/68 37/57 43/54 39/57 45/54 49 44662 < 45363 233 59/48 49/60 62/56 60/51 50 45371 > 46594 407 p47 58/58 60/54 58/57 50/60 55/57 68/60 61/60 63/56 74/53 61/57 56/63 51/53 79/54 58/53 78/52 51 46629 > 47339 236 BV-e31 59/63 61/60 59/58 51/63 56/58 69/67 62/67 65/57 77/56 62/64 57/69 52/66 82/56 59/57 79/57 52 47357 > 47809 150 p24 60/48 62/47 60/47 52/56 57/47 71/52 63/53 66/47 78/40 63/48 58/51 53/47 83/39 60/48 80/40 53 47873 < 48184 103 38.7k 63/36 54/33 65/33 67/33 65/34 59/41 54/43 54 48165 < 48953 262 lef-1 62/51 64/48 63/59 55/59 60/58 74/56 66/59 68/53 80/45 66/51 60/56 55/47 85/47 62/48 82/45 55 48865 > 50529 554 pif-1 63/46 65/37 64/47 56/50 61/46 75/50 67/50 69/44 82/41 67/45 61/51 7/42 87/40 64/41 84/40 56 50540 < 51262 240 fgf-1 64/39 66/31 65/41 57/43 62/42 76/39 68/39 70/32 83/29 69/46 62/45 56/42 88/31 66/32 85/30 57 51944 < 53362 472 bro-A 54/67 137/70 111/74 60/71 58 53820 < 54143 107 U 59 54398 > 54784 128 65/42 67/37 67/39 59/36 64/42 79/35 70/36 73/32 85/31 71/37 64/44 59/65 90/29 68/47 87/31 60 54803 < 55120 105 lef-6 67/42 68/31 68/29 60/38 65/28 80/39 71/40 74/33 86/30 72/35 65/43 60/32 91/32 69/39 88/34 61 55162 < 56094 310 dbp 68/29 69/26 69/30 61/31 66/29 81/33 72/29 75/25 73/31 66/27 61/27 70/29 62 56108 < 56377 89 62/38 82/36 73/40 63 56371 > 56559 62 dnapol 64 56636 < 56962 108 70/37 63/31 67/38 82/38 73/34 77/35 67/45 65 57010 > 58194 394 p45/p48 69/66 72/61 71/64 64/71 68/64 83/72 74/72 78/57 90/55 75/67 68/73 63/51 95/55 73/52 91/55 66 58221 > 58595 124 P12 70/57 73/48 72/59 65/53 69/56 84/55 75/54 79/43 91/44 76/32 69/60 64/38 96/44 74/42 92/44 67 58632 > 59789 385 p40/c42 71/56 74/51 73/58 66/58 70/58 85/58 76/59 80/49 92/46 77/49 70/60 66/44 97/46 75/45 93/46 68 59805 > 59978 57 p6.9a 72/ 75/86 74/63 67/ 71/63 86/60 77/ 81/46 93/71 78/ 71/56 67/57 98/86 94/71 69 60065 < 60838 257 lef-5 73/55 76/51 75/61 68/61 72/61 87/59 78/59 82/51 94/51 79/59 72/65 69/52 99/48 76/50 95/51 70 60791 > 61777 328 38k 74/56 77/49 76/62 69/58 73/62 88/58 79/56 83/49 95/47 80/57 73/63 70/46 100/49 77/50 96/46 71 61737 < 62075 112 pif-4 75/50 78/50 77/49 70/54 74/51 89/51 80/54 84/45 96/44 81/41 74/56 71/42 101/45 79/48 97/44 72 62210 > 65695 1161 helicase 76/38 79/32 78/38 71/40 75/38 90/37 81/36 85/30 97/31 82/32 75/41 72/32 102/31 80/31 98/31 73 65828 < 66475 215 odv-e25 77/63 81/56 79/56 72/64 76/56 91/66 82/67 86/60 98/56 83/60 76/65 74/60 103/55 81/57 99/56 74 66599 < 67078 159 p18 78/39 82/45 80/50 73/55 77/50 92/55 83/49 87/50 99/46 84/49 77/63 75/43 104/46 82/39 100/48 75 67148 > 67939 263 p33 79/53 83/53 81/55 74/58 78/54 93/57 84/56 88/57 100/52 85/55 78/66 76/55 105/52 83/51 101/51 76 68076 < 69524 482 lef-4 80/48 85/45 83/46 75/53 80/46 95/47 86/46 91/47 112/41 87/50 80/54 78/42 114/41 86/44 110/42 77 69615 > 70508 297 vp39 81/61 86/33 84/60 76/58 81/59 96/52 87/53 92/39 113/36 88/48 81/65 79/33 115/36 87/36 111/35 78 70586 > 71488 300 odv-e27 82/51 87/41 85/57 77/55 82/58 97/55 88/53 93/41 114/40 89/42 82/60 80/36 116/41 88/43 112/41 79 71520 > 71729 69 U 80 72155 < 72967 270 83/30 88/24 86/29 78/35 83/29 99/27 90/30 94/24 116/27 92/27 83/28 82/31 117/26 89/26 113/27 81 73024 > 73302 92 U 82 73324 < 75132 602 vp91 85/40 91/32 88/40 80/35 86/39 101/40 92/41 96/34 121/39 94/42 85/40 84/35 122/36 92/33 118/40 83 75104 > 75601 165 tlp20 86/30 84 75618 < 76010 130 U 85 76616 > 78076 486 bro-B 101/66 167/59 109/65 86 78318 > 78800 160 bro-C 110/48 111/44 108/46 87 79019 > 79636 205 87/64 94/55 90/64 82/71 88/69 103/73 94/72 98/67 123/57 96/69 87/70 86/64 124/57 94/57 120/57 88 79596 > 80468 290 gp41 88/46 95/55 91/57 83/61 89/59 104/66 95/63 99/59 124/47 97/58 88/63 87/44 125/48 95/53 121/48 89 80491 > 80805 104 90/41 85/40 90/35 105/39 96/46 98/40 89/42 90 80771 > 81895 374 vlf-1 91/67 97/61 93/67 86/67 91/67 106/71 97/69 101/48 126/53 99/68 90/76 89/49 127/51 97/55 123/53 91 81912 > 82166 84 92/68 99/58 94/37 88/76 92/70 107/70 98/70 102/62 128/56 100/64 91/81 91/56 129/56 99/58 125/54 92 82800 > 83660 286 109/31 117/27 111/29 106/34 110/29 130/35 118/39 121/26 109/30 108/29 119/30 93 83938 > 84510 190 rr1 127/26 119/32 94 84571 < 85893 440 alk-exo 107/38 115/38 110/40 104/40 109/40 125/39 115/38 119-120/40 146/38 114/39 107/41 106/36 152/39 117/33 145/38 95 85955 < 86251 98 106/40 109/49 103/62 108/48 124/55 114/54 115/53 106/45 96 86359 > 87711 450 fgf-2 105/26 113/31 108/31 102/43 107/32 123/34 113/38 118/32 145/25 116/40 105/36 104/29 151/25 116/28 144/27 97 88307 < 88672 121 U 98 88812 > 90563 583 ligase 103/49 110/50 105/48 99/55 104/49 120/54 110/56 115/46 142/43 112/50 102/61 101/42 148/43 110/44 141/43 99 90706 < 91161 151 fp 25k 101/50 108/56 103/57 97/57 102/56 118/56 108/58 113/49 141/51 110/53 100/55 100/48 146/44 109/51 140/51 100 91234 < 92727 497 lef-9 100/68 107/65 102/64 96/69 101/66 117/68 107/71 112/69 140/62 109/73 99/72 99/64 145/62 107/65 139/61 101 93356 < 94207 283 iap-5 99/49 106/43 101/54 95/54 100/53 116/53 106/53 111/44 139/34 108/54 98/58 98/36 143/33 106/38 137/34 102 94261 < 94767 168 98/31 105/25 101/33 94/32 100/34 115/26 105/24 110/28 107/30 97/31 105/47 103 94773 < 95159 128 pif-6 97/59 104/54 100/56 93/65 98/56 114/60 104/60 109/48 137/45 106/51 96/57 96/51 141/48 104/48 135/45 104 95125 > 96204 359 lef-3 96/25 99/23 92/24 97/25 113/27 108/23 95/24 105 96201 < 96500 99 69/43 74/42 75/43 106 96596 < 98482 628 desmoplakin 95/28 102/26 98/23 96/26 112/30 102/26 107/32 104/27 94/31 107 98481 > 101741 1086 polymerase 94/48 101/46 97/48 90/48 95/47 111/49 101/47 106/45 134/44 103/47 93/49 93/40 138/43 101/41 132/44 108 101738 < 102205 155 93/56 100/48 96/47 89/57 94/48 108/53 99/52 103/46 129/28 101/55 92/52 92/41 130/28 100/40 126/28 109 102606 < 105233 875 lef-8 110/66 118/62 112/61 107/67 111/62 131/67 119/67 122/63 149/61 121/67 110/68 109/61 155/61 121/60 148/61 110 105394 > 105786 130 113/26 113/26 111/28 111 105857 < 106063 68 108/35 114/32 133/40 168/31 112/50 172/30 170/29 112 106065 > 106466 133 111/56 122/51 115/54 109/59 115/55 134/60 121/62 125/52 169/41 122/63 113/64 112/46 173/40 127/39 171/41 113 106450 < 107439 329 113/40 124/38 116/31 110/30 116/30 135/30 122/33 126/37 170/33 123/37 114/32 174/28 128/37 172/33 114 107466 < 107660 64 114/48 117/42 111/44 117/45 136/35 123/35 115/55 115 107738 > 108745 335 vp1054 115/50 127/46 119/54 113/54 119/55 138/52 125/54 129/44 173/41 126/48 116/56 115/39 177/43 130/41 175/42 116 108764 > 108988 74 U 117 108999 > 110159 386 fgf-3 117/36 128/38 121/40 114/35 121/35 140/38 127/40 131/30 128/32 118/42 117/27 133/28 118 110278 > 111231 317 me-53 119/46 131/39 123/47 116/51 123/48 143/50 129/51 133/32 178/33 130/46 120/55 120/40 182/32 134/34 180/32 Note: The viruses used in this table are Adoxophyes orana granulovirus (AdorGV), Agrotis segetum granulovirus (AgseGV), Clostera anastomosis granulovirus (CaLGV), Choristoneura occidentalis granulovirus (ChocGV), Clostera anachoreta granulovirus (ClanGV), Cydia pomonella granulovirus (CpGV), Cryptophlebia leucotreta granulovirus (CrleGV), Epinotia aporema granulovirus (EpapGV), Helicoverpa armigera granulovirus (HearGV), Phthorimaea operculella granulovirus (PhopGV), Pieris rapae granulovirus (PrGV), Plutella xylostella granulovirus (PlxyGV), Pseudaletia unipuncta granulovirus (PsunGV), Spodoptera litura granulovirus (SpltGV) and Xestia c-nigrum granulovirus (XecnGV).
a: This ORF has only low identity to several NPVs;
U: These ORFs are unique to CnmeGV genomeTable S1. Analysis and homology search of CnmeGV ORFs

 DownLoad:
DownLoad: