












-
Dear Editor,
Viruses are the most abundant, diverse, and ubiquitous entities(approximately 1031)on Earth. They play major roles in horizontal gene transfer, the regulation of bacterial community structures, as well as nutrient and energy cycles of marine ecosystems(Danovaro et al., 2008). In particular, lytic bacteriophages(phages)can infect and kill bacteria without harming human or animal hosts, meaning that their influence on bacterial communities is extremely focused and direct(Xu et al., 2015). Thus, lytic phages are studied in a wide variety of ecosystems(e.g., marine, lake, and polar environments)(Danovaro et al., 2008); unfortunately, knowledge of phages in wetland s is scant. wetlands are considered to be the most biologically diverse of all ecosystems and comprise 6% of Earth's surface. Often called the "kidney of the Earth, " wetland s are not only important reservoirs of biodiversity, but also participate in flood control, water purification, and climate change adaptation. However, wetland s are fragile and easily disrupted. Thus, the investigation of lytic phage diversity in wetlands will increase our understanding of their effects on the microbial community. Our results should contribute to conservation efforts and the maintenance of the delicate ecological balance in these ecosystems.
Napahai is a typical degraded plateau wetland, with a low latitude, high altitude, and low temperature(annual mean temperature of 5.4 ℃). The lytic bacteriophages here tend to be cold-active, proliferating at temperatures ≤ 4 ℃. In this letter, the morphological diversity of cultured cold-active lytic bacteriophages, sampled from Napahai, was systematically examined using transmission electron microscopy(TEM), a common and reliable technique for classifying phages.
Water and soil samples were obtained from the Napahai plateau wetland (E99°37′20″-E99°38′16″, N27°50′01″-N27°54′26″; altitude 3263-3280 m, average temperature 4-7 ℃, and pH 5.5-7.0)located in Northwest Yunnan province, China. A 2-kg soil sample(from a depth of 10 cm) and a 5-L water sample(surface water)were collected in September 2014. For culturing, we chose the eutrophic Luria-Bertani(LB)medium and the oligotrophic Peptone-Yeast powder-Glucose-Vitamins(PYGV)medium; these media were considered ideal for isolating cold-adapted phages from the Napahai wetland in a previous study(Ji et al., 2015). Water samples were filtered through a 0.22-µm filter(Millipore Corp., Bedford, USA) and soil samples were dissolved in ddH2O(1 g soil per 50 mL water). They were then spread onto LB and PYGV solid plates and incubated for 2-3 days at 15 ℃.
Approximately 37 bacterial isolates were obtained based on color, Gram staining, and morphology. These bacteria were used as host cells to isolate and propagate phages. We enriched the phages by pre-incubating filtered water(50 mL through 0.22-µm filters) and dissolved soil(1 g per 50 mL of liquid LB and PYGV)samples with the 37 host bacteria. Samples were subjected to static liquid incubation at 15 ℃ for 10 days(Ji et al., 2015) and then centrifuged(12000 g, 15 min, 4 ℃, Beckman Avanti J-25, USA). The supernatants were filtered through 0.22-µm filters(Millipore Corp., Bedford, USA).
Eighteen bacterial isolates were sensitive to phage infection. Molecular identification of isolates was confirmed through 16S rRNA gene sequencing with universal PCR primers(Ji et al., 2015). The nucleotide sequences of host cells have been deposited in NCBI(accession numbers KM391403 to KM391420). Based on the 16S rRNA gene sequence analysis, the 18 isolates were grouped into four different classes(Actinobacteria, Flavobacteria, Bacilli, and Gamma-Proteobacteria; Supplementary Table S1), and a phylogenetic tree was constructed using the neighbor-joining method in MEGA 6.0 with 1000-fold bootstrap support(Supplementary Figure S1). All sequences corresponded to known bacteria genera. Nine sequences fell into the cluster corresponding to Pseudomonas spp., three to Flavobacterium spp., and one respectively to Acinetobacter spp., Stenotrophomonas spp., Arthrobacter spp., Bacillus spp., Aeromonas spp., and Exiguobacterium spp.. The target bacterial cell morphotype(rod-shaped cells)dominated the bacterial community in the Napahai wetland (Supplementary Table S1), consistent with a report from Lake Créteil, a eutrophic ecosystem(Ram et al., 2010). Previous studies have found that higher percentages of rod-shaped cells contribute to an increased bacterial carbon pool in the Northern Adriatic Sea, and rod-shaped cells appear to favor nutrient-rich environments(Ferla and Leonardi, 2005). Our results, coupled with previous data, suggest that the trophic conditions of the Napahai plateau wetland have changed to eutrophic conditions; thus, some measures should be taken to protect the wetland from further eutrophication.
Phages were isolated and purified through successive single-plaque isolation with the double-layer agar method(Xiang et al., 2005). Each phage was pelleted using PEG 8000 and CsCl ultracentrifugation(Ji et al., 2015). Eighteen phages showed clear plaques after incubation at 4 ℃ for 48 h on LB or PYGV double-layer agar plates, indicating they are likely cold-active lytic phages(Supplementary Table S2). Cold-active phages have been isolated from both natural and anthropogenic low-temperature environments, including oceans, glaciers, sewage, and refrigerated food storage(Sawstrom et al., 2008). The cold-active phages in this study were lytic only against their own hosts and therefore showed high specificity for host cells(Supplementary Table S3).
We examined key morphological features(capsid diameter, tail length, and tail dimensions)of purified phages using TEM of negatively stained preparations with 2%(w/v)uranyl acetate(Supplementary Table S4). We isolated six Myoviridae, four Podoviridae, and eight Siphoviridae, based on descriptions from the International Committee on Taxonomy of Viruses(Figure 1). Although all of the phage particles observed belonged to tailed phages and had icosahedral heads, they varied in head diameters, tail lengths, and other tail dimensions(Supplementary Table S4). These data demonstrate that the phages in the Napahai wetland s are fairly diverse.
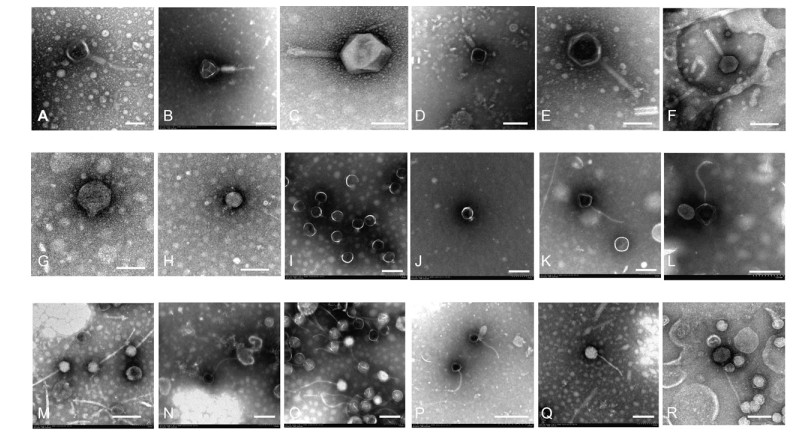
Figure 1. Transmission electron micrographs of phages isolated from the Napahai plateau wetland, demonstrating the diversity of viral morphotypes. A-F, Myoviridae possessing contracted (A-D) tail sheath with central tube particles and (E-F) non-contracted tail sheath. G-J, Podoviridae. K-R, Siphoviridae. Scale bars represent 100 nm except I (50 nm) and P (200 nm).
In aquatic environments, ≥ 65% viruses have capsid diameters of 30-60 nm(Wommack and Colwell, 2000). Viruses smaller than 60 nm(68%)were dominant at 7 sites in the West African Inland Aquatic Systems(Bettarel et al., 2006) and accounted for 75% of all observed phages(diameters between 20-50 nm)within Chesapeake Bay sediments(Helton et al., 2012). Viral capsids also usually range in size from 30 to 60 nm in temperate freshwater environments(Williamson et al., 2005). In contrast to these previous reports, the capsid diameters of the Napahai phages tended to be larger, with 56% of all observed phages having diameters over 60 nm and 22% having diameters over 100 nm(mainly Myoviridae). Our results are similar to findings from Mono Lake, where 40% of the viruses had capsid diameters of 30-60 nm and the majority had capsids larger than 60 nm, but none had capsid diameters ≥ 110 nm(Brum and Steward, 2010). The capsid size distributions we observed may be a consequence of the specific hydrographic and geological conditions in the Napahai region.
Tailed phages belong to the order Caudovirales, which all possess double-stranded DNA genomes. Most phages in aquatic environments are Caudovirales, including Myoviridae, Siphoviridae, and Podoviridae, with Siphoviridae thought to be the most abundant morphotype. Indeed, 61% Siphoviridae, 24% Myoviridae, 14% Podoviridae, and 1% non-tailed phages were found among 5500 isolates from diverse aquatic ecosystems(Ackermann, 2007). Moreover, 50% Siphoviridae, 30% Myoviridae, and 19% Podoviridae were reported in a lake environment(Demuth et al., 1993). In Mono Lake, Siphoviridae was the dominant morphotype(Brum and Steward, 2010). Although culture collections may exhibit bias, we were able to isolate eight Siphoviridae phages among 18 cold-active lytic isolates.
In summary, rod-shaped bacterial cells and Pseudomonas spp. dominated the bacterial community in the Napahai plateau wetland. Based on our TEM results, bacteriophages with larger capsids(60-140 nm) and Siphoviridae members were dominant. We were able to isolate a batch of cultured cold-active bacteriophages and their host bacterial strains, which should provide opportunities for further qualitative or quantitative analysis of bacteriophage function in wetland ecology. Specifically, these new isolates can help clarify bacteriophage cold-adaptation mechanisms, structure, and assembly, as well as improve our understanding of phage-host co-evolution and interactions.
HTML
-
This work was supported by the National Natural Science Foundation of China(31160121, 31360129) and the China Scholarship Council. This article does not contain any studies with human or animal subjects performed by any of the authors. Further, the authors declare that they have no conflict of interest.
Supplementary Figures/Tables are available on the website of Virologica Sinica: www.virosin.org; link.springer.com/journal/ 12250.
-
Phylum Class Order Family Genus Gram staining Morphology PhageFamily Host trains Actinobacteria Actinobacteria Actinomycetales Micrococcaceae Arthrobacter G+ rod Siphovirus LSW-9 Bacteroidetes Flavobacteria Flavobacteriales Flavobacteriaceae Flavobacterium G- rod Siphovirus PSW-7, PSW-12 Proteobacteria Gammaproteobacteria Pseudomonadales Pseudomonadaceae Pseudomonas G- rod Siphovirus LS-15; LS-4; PS-7; PSW-10 Firmicutes Bacilli Bacillales Bacillaceae Exiguobacterium G+ rod Siphovirus LSW-8 Firmicutes Bacilli Bacillales Bacillaceae Bacillus G+ rod Podoviridae LS-8 Proteobacteria Gammaproteobacteria Pseudomonadales Moraxellaceae Acinetobacter G- rod Podoviridae LSW-7 Proteobacteria Gammaproteobacteria Pseudomonadales Pseudomonadaceae Pseudomonas G- rod Podoviridae PS-1; PSW-5 Proteobacteria Gammaproteobacteria Pseudomonadales Pseudomonadaceae Pseudomonas G- rod Myoviridae PS-3, PS-4, PS-12; Proteobacteria Gammaproteobacteria Xanthomonadales Xanthomonadaceae Stenotrophomonas G- rod Myoviridae LSW-10 Proteobacteria Gammaproteobacteria Aeromonadales Aeromonadaceae Aeromonas G- rod Myoviridae NPH-1 Bacteroidetes Flavobacteria Flavobacteriales Flavobacteriaceae Flavobacterium G- rod Myoviridae PSW-8 Table S1. Host isolate and phage taxonomy
Figure S1. Neighbor-joining phylogenetic tree based on 16S rRNA gene sequences available from the NCBI GenBank
Phage 4℃ 10℃ 15℃ 20℃ 25℃ 30℃ 37℃ 42℃ LS-4 + + + + + + - - LS-8 + + + + + + + - LS-15 + + + + - - - - PS-1 + + + + - - - - PS-3 + + + + + - - - PS-4 + + + + - - - - PS-7 + + + + - - - - PS-12 + + + + + - - - NPH-1 + + + + - - - - LSW-7 + + + + + - - - LSW-8 + + + + + + + - LSW-9 + + + + + - - - LSW-10 + + + + - - - - PSW-5 + + + + - - - - PSW-7 + + + + + - - - PSW-8 + + + + + - - - PSW-10 + + + + - - - - PSW-12 + + + + + - - - Note: "+"indicates plaque formation, "-" indicates no plaque formation. Table S2. Temperature ranges of plaque formation

LS-4 LS-8 LS-15 PS-1 PS-3 PS-4 PS-7 PS-12 NPH-1 LSW-7 LSW-8 LSW-9 LSW-10 PSW-5 PSW-7 PSW-8 PSW-10 PSW-12 VLS-4 + - - - - - - - - - - - - - - - - - VLS-8 - + - - - - - - - - - - - - - - - - VLS-15 - - + - - - - - - - - - - - - - - - VPS-1 - - - + - - - - - - - - - - - - - - VPS-3 - - - - + - - - - - - - - - - - - - VPS-4 - - - - - + - - - - - - - - - - - - VPS-7 - - - - - - + - - - - - - - - - - - VPS-12 - - - - - - - + - - - - - - - - - - VNPH-1 - - - - - - - - + - - - - - - - - - VLSW-7 - - - - - - - - - + - - - - - - - - VLSW-8 - - - - - - - - - - + - - - - - - - VLSW-9 - - - - - - - - - - - + - - - - - - VLSW-10 - - - - - - - - - - - - + - - - - - VPSW-5 - - - - - - - - - - - - - + - - - - VPSW-7 - - - - - - - - - - - - - - + - - - VPSW-8 - - - - - - - - - - - - - - - + - - VPSW-10 - - - - - - - - - - - - - - - - + - VPSW-12 - - - - - - - - - - - - - - - - - + Note: "+"indicates could infect host cell, "-" indicates could not infect host cell. Table S3. The host ranges of isolated phages
Number Family Head capsid diameter (nm) Tail length (nm) Tail diameter (nm) Host strain A Myoviridae 116.0±2.31 296.7±3.01 29.3±0.76 PS-4 B Myoviridae 108.6±1.86 173.3±2.55 32.4±1.22 LSW-10 C Myoviridae 134.1±1.56 154±3.02 38.2±1.15 PSW-8 D Myoviridae 70.9±2.09 106.5±2.63 25.8±0.56 PS-3 E Myoviridae 121.6±2.47 194.6±2.17 21.6±1.09 NPH-1 F Myoviridae 70.3±0.98 118.9±1.83 20.5±0.66 PS-12 G Podoviridae 60.8±1.10 13.5±0.58 10.8±0.87 LSW-7 H Podoviridae 78.0±1.36 28.2±0.72 11.4±1.20 PS-1 I Podoviridae 57.0±1.53 23.3±0.68 13.6±1.27 PSW-5 J Podoviridae 57.7±1.64 15.4±0.71 7.7±0.88 LS-8 K Siphoviridae 76.9±2.31 192.3±1.93 7.7±0.85 PSW-10 L Siphoviridae 52.7±2.01 196.2±1.84 8.1±1.10 PS-7 M Siphoviridae 57.7±1.86 199.6±1.76 11.5±0.78 LSW-8 N Siphoviridae 53.3±1.42 166.7±1.68 6.7±0.34 LSW-9 O Siphoviridae 58.7±1.38 204.2±2.04 7.6±0.48 LS-15 P Siphoviridae 59.1±2.04 190.1±1.57 9.1±0.43 PSW-7 Q Siphoviridae 42.3±1.67 165.4±1.36 7.7±0.50 PSW-12 R Siphoviridae 83.3±2.34 190.5±3.05 9.5±0.62 LS-4 Note: Means ± SD (n = 3) Table S4. Mean sizes of phages observed by TEM isolated from the Napahai plateau wetland, Yunnan province, China

 DownLoad:
DownLoad: