












-
Dear Editor,
Influenza A viruses (IAVs) are single-stranded, negative sense RNA viruses. IAV subtype is determined on the basis of the viral surface glycoproteins, hemagglutinin (HA), and neuraminidase (NA). To date, 18 HA and 11 NA subtypes have been reported (Tong et al., 2012). IAVs can cause sporadic infections, local epidemics, and global pandemics among humans. In addition to humans, IAVs can naturally infect avian, swine, equines, canines, and sea mammals(Webster et al., 1992). Migratory waterfowl are the natural reservoir for IAVs, and the avian-origin IAVs play an important role in influenza ecology and have been involved in generation of the IAVs infection in humans. At least one or more genetic segments of all four known pandemic strains are of avian origin, and these avian-origin genes reassorted with those IAVs from domestic animals to generate pandemic viruses. For example, the HA genes (major antigenic determinants) of 1918, 1957, and 1968 pandemic viruses are all of avian-origin (Webster et al., 1997); the 2009 H1N1 pandemic virus has avian-origin PB2 and PA genes (Shinde et al., 2009). Besides pandemic viruses, in the past decades, there have also been a number of reported human infections with avian IAVs, including subtypes H5N1, H6N1, H7N2, H7N3, H7N7, H9N2, H10N7 and H7N9. Thus, monitoring the evolution of avian IAVs and rapidly detecting these viruses in human are important components of influenza surveillance and pandemic preparedness.
On November 27, 2013, a novel H10N8 IAV was recovered from a patient with fatal pneumonia in Nanchang, Jiangxi Province, China (Chen et al., 2014). This virus had a Eurasian-lineage H10 gene and a North American-lineage N8 gene. Two more human cases of H10N8 virus infection were reported in the same city in the next few months. Recent serologic surveillance in this region showed that birds, including chicken and ducks, from five live bird markets (LBM) were seropositive for H10 IAV, indicating that H10 virus has spread widely in the region, posing a potential threat to public health (Xu et al., 2015).
Quantitative RT-PCR (qRT-PCR) has been commonly used for IAV detection in influenza surveillance and disease diagnosis because of its high sensitivity, specificity, and high throughput. A number of HA subtype specific qRT-PCR methods have been developed, including H5, H7, and H9 (Monne et al., 2008). However, there is still lack of a specific qRT-PCR method for detecting the emerging H10N8 IAVs. In this study, a Eurasian-lineage H10 specific qRT-PCR is developed and validated.
To develop Eurasian-lineage H10 gene specific probe, a total of 1, 090 available H10 genes were retrieved from the Influenza Virus Resource (http://www.ncbi.nlm.nih.gov/genomes/FLU/FLU.html), including 470 Eurasian-lineage H10 genes and 620 North American-lineage H10 genes. The nucleotide sequences were aligned using MUSCLE (Edgar, 2004), and a Eurasian-lineage H10 specific primer set and Taqman probe were designed using Primer Express version 2.0 (Applied Biosystems Inc., Foster City, CA, USA): forward primer (H10F) 5′-AATTTTGTTCC[A/G]GTTGTTGG[A/G]GCA-3′, reverse primer (H10R) 5′-TTTGCTAACTCG[A/G]CT[T/C]GG[T/C]GCTAT-3′ (Eurofins Genomics, Huntsville, Alabama, USA); the MGB-labeled probe FAM-5′-CCTCAGGTCAATGGACAAAG-3′-MGB-NFQ targeting the region covering 683–826 (144 bp total) (Thermo Fisher Scientific, Grand Island, NY, USA). Sequence analyses showed that, at the region (from nucleotide position 710 to 729) covered by probe, there was a three-nucleotide discrepancy (marked in bold) between the H10 IAVs of Eurasian-lineage (CCTCAGGTCAATGGACAAAG) and those of North American-lineage (CCACAGG[A/G]CAATGG[C/T]CAAAG). Thus, this probe was proved to be Eurasian-lineage H10 gene specific.
The qRT-PCR method was performed in a 20-μL volume containing 5 μL of 4 × TaqMan Fast Virus 1-Step Master Mix (Thermo Fisher Scientific, Grand Island, NY, USA), 1 μL of forward primer H10F (900 nmol/L), 1 μL of reverse primer H10R (900 nmol/L), 1 μL of probe (100 nmol/L), 1 μL viral RNA (30 ng) being tested, and 11 μL DEPC-treated water. The qRT-PCR was performed using the Mx3000P qPCR System (Stratagene, La Jolla, CA, USA) with the following cycling conditions: reverse transcription at 50 ℃ for 5 min and 95 ℃ for 20 sec, followed by 40 cycles of PCR at 95℃ for 3 sec and 60℃ for 30 sec.
Using the reassortant virus A/chicken/Jiangxi/JXA132717/2013 (HA, NA) × A/Puerto Rico/8/1934 (H10N8), the sensitivity assays showed that the H10-specific qRT-PCR method detected a viral load as little as 2.32 TCID50/mL with a Ct value of 31.38±0.17 (Table 1).
Dilution TCID50/mL Average Ctvalue±standard deviationa 10−1 2.32 × 103 21.34±0.07 10−2 2.32 × 102 24.53±0.10 10−3 2.32 × 101 28.12±0.39 10−4 2.32 × 100 31.38±0.17 10−5 2.32 × 10−1 39.51±0.21 10−6 2.32 × 10−2 No Ct 10−7 2.32 × 10−3 No Ct 10−8 2.32 × 10−4 No Ct Negative controlb - No Ct Note:a: The Ct values were calculated based on the dRn threshold of 0.27 adapted from (Ben Shabat et al., 2010); b: DEPC treated water. Table 1. Detection limits of viral load for a Eurasian-lineage H10 specific quantitative RT-PCR method
To quantify the copies of viral genes, a plasmid containing 9.4×102 to 9.4×107 copies H10 gene of A/chicken/Jiangxi/JXA132717/2013 (H10N8) was used to establish a standard curve. Standard curves for those plasmids were generated by plotting their cycle threshold numbers (Ct) versus their dilution factors. The resulting formula was log10 (x copy number of H10 genes)=-3.699 (ΔCt) + 20.570 (R2=0.998, p < 0.001), which was used to measure the minimum copy number of H10 genes to be detected by qRT-PCR. Results showed that the H10-specific qRT-PCR method detected as few as 76 copies of H10 genes.
To assess specificity of the proposed qRT-PCR method, the specificity assays included 23 isolates of H1, H2, H3, H4, H5, H6, H7, H9 IAVs, one H10 reassortant virus containing Eurasian-lineage H10 gene, and 2 North American-lineage H10 isolates, as well as four other viruses commonly reported in domestic poultry, including infectious bursal disease virus, infectious bronchitis virus, Newcastle disease virus, and reovirus (Supplementary Table 1). The specificity analyses demonstrated that the proposed qRT-PCR method did not react with the two testing North America-lineage H10 IAVs, other subtype IAVs, or other avian viruses (Supplementary Figure 1), indicating this method is highly specific for Eurasian-lineage H10 IAVs.
This method was then applied in identifying viruses from 49 cloaca swab samples from a LBM in Nanchang city, China and another 50 cloaca swab samples collected from migratory waterfowl from the Mississippi migratory bird flyway. All these 99 samples were shown to be positive for IAV RNA using a qRT-PCR matrix gene assay. Of these 49 samples from Nanchang, 15 were cloacal swab samples from waterfowl species, including ducks and geese; 30 were oropharyngeal or cloacal swab samples from terrestrial birds, including chickens, pigeons, guinea fowl, and turtle doves; and 4 were feces samples from the environment. Among the 49 samples from Nanchang, 44 were described in previous study and 30 were successfully sequenced samples included 18 H10, 2 H6, and 10 H9 subtypes of IAVs. Results from this study showed that, in addition to those H10 reported before (Xu et al., 2015), 7 more samples from Nanchang were identified as H10 positive. Taken together, 25 (51.0%) of 49 samples from Nanchang were H10 positive, as were confirmed by genomic sequencing (Eurofins MWG Operon Inc., Huntsville, AL, USA). The HA gene of two newly identified H10 viruses were sequenced with GenBank accession numbers of KU513668 and KU513669. No H10 positive samples were detected from those from Mississippi migratory bird flyway.
To evaluate the genetic relations of newly identified H10 genes with those reported before, phylogenetic analyses were performed using maximum likelihood in RAxMLv7.6.8 (Stamatakis, 2006), and a general time-reversible nucleotide substitution model and gamma model of rate heterogeneity were used. From the 1, 090 available H10 genes from public databases (accessed on August 3, 2015), 144 representative H10 sequences were used for final phylogenetic analyses. The selection of viral sequences attempted to maximize the coverage of temporal and spatial distribution, and the branches on the tree tropology for the viruses. Phylogenetic analyses showed those H10 genes detected formed a monophyletic Eurasian clade together with the HA gene of human H10N8 virus A/Jiangxi-Donghu/346/2013 (H10N8) (Figure 1).

Figure 1. Phylogenetic analysis of HA genes of H10N8 viruses from a live bird market in Nanchang city, China. The viruses marked in blue indicate those detected previously (Xu et al., 2015); the viruses marked in red indicate those newly detected by qRT-PCR in this study; the human origin A/Jiangxi-Donghu/346/2013 (H10N8) virus was marked in purple. GenBank accession numbers are included at the end of each virus name. Jiangxi is abbreviated as JX. Host species are: Ck (chicken), Mdk (migratory duck), and Dk (duck).
The probe used in this study was designed to target a highly conserved region in the H10 genes of Eurasian lineage, which had a three-nucleotide discrepancy from those of North American lineage. To enhance the probe specificity, we incorporated TaqMan probe with a non-fluorescent MGB quencher, which can improve signal-to-noise ratios (Afonina et al., 2002). MGB could increase the melting temperature about 10℃ and help discriminate mismatched hybridization, and it could also enhance sensitivity of the probe over that of a conventional TaqMan probe (Lew et al., 2004).
Contrasted to classical diagnosis method such as influenza virus isolation in MDCK or embryonated eggs, RT-PCR could generate results within a single working day, greatly shortening the time of detection (Fouchier et al., 2000). Notably, our method allowed for high input of samples and avoided cross contamination by eliminating the post reverse transcription steps that are required by conventional RT-PCR.
Intercontinental gene flow through the migration of wild birds was documented in a few occasions. The introduction of Eurasian-lineage H6 viruses has resulted in the replacement of the endemic avian influenza viruses in North America (Lam et al., 2012). Recently, migration of wild birds has led to the spread of H5 highly pathogenic avian influenza virus from Eurasia to North America, and it has consequently caused outbreak in domestic poultry in the United States (Ip et al., 2015). Influenza surveillance in the Mississippi migratory bird flyway (from July 2010 to January 2011) detected a Eurasian-lineage H10 IAVs in North America (Fries et al., 2014). In this study, we applied our method in a small size of samples from North America, and no Eurasian-lineage H10 positive samples were detected. Nevertheless, this method can be used for detection of Eurasian-lineage H10 IAVs in North America.
In conclusion, the TaqMan-MGB qRT-PCR developed in this study can be used in influenza surveillance for rapidly detecting Eurasian-lineage H10 IAVs.
HTML
-
This study was partially supported by the National Institutes of Health (grant no. P20GM103646) and the United States Department of Agriculture Animal and Plant Health Inspection Service (agreement 14-7428-1041-CA). The authors declare no interests of conflicts. The sample collection in animals was compliant with the protocol (no. 13-048) approved by the Institutional Animal Care and Use Committee, Mississippi State University.
Supplementary table/figure are available on the website of Virologica Sinica: www.virosin.org; link.springer.com/journal/12250.
-
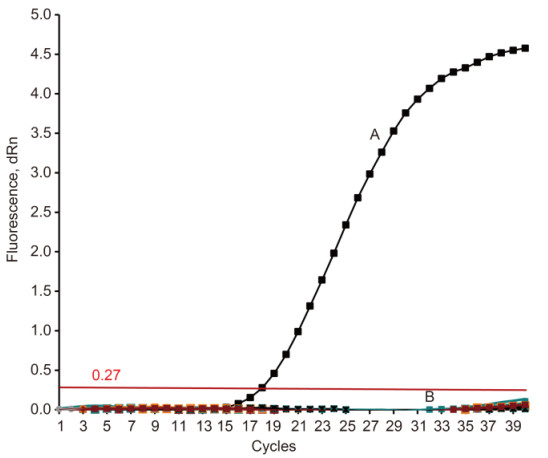
Figure S1. Specificity of quantitative RT-PCR for the detection of Eurasian-lineage H10 influenza A virus. Line A represents the amplification for the reassortant virus A/chicken/Jiangxi/JXA132717/2013 (HA, NA) × A/Puerto Rico/8/1934 (H10N8) amplification; Line B represents the amplifications of other influenza virus subtypes, including H1, H5, H7, H9, and North American-lineage H10N7 viruses, as well as other non-influenza avian viruses (Newcastle disease virus, infectious bursal disease virus, infectious bronchitis virus, and reovirus). The control reaction was conducted by using DEPC-treated water as template. The number of cycles (Ct) was calculated based on the maximum baseline-corrected normalized reporter (dRn) threshold of 0.27. Each assay was performed in triplicate.
Host Virus name Human A/Puerto Rico/8/1934(H1N1) A/California/04/2009(H1N1) A/Brisbane/10/2005(H3N2) A/Perth/16/2009(H3N2) Avian A/mallard/Oregon/A00544081/2008(H1N1) A/mallard/Wisconsin/A00751454/2009(H1N1) A/mallard/Oregon/A00300758/2008(H2N3) A/mallard/Oregon/A00320372/2007(H2N3) A/mallard/Wisconsin/A00661712/2009(H3N2) A/mallard/Wisconsin/A00714818/2009(H3N2) A/mallard/Colorado/A00170379/2006(H3N8) A/mallard/Wisconsin/A00170366/2006(H3N8) A/mallard/Washington/A00714770/2009(H4N6) A/mallard/Washington/A00757090/2009(H4N6) A/chicken/Jiangxi/JXA132321/2013(H5N?)a A/mallard/Colorado/A00551322/2007(H6N1) A/mallard/New York/A00603000/2009(H6N1) A/blue-winged teal/Illinois/10OS3610/2010(H6N2) A/mallard/Ohio/81/1986(H6N2) A/mute swan/Rhode Island/A00325114/2008(H7N3) A/american green-winged teal/Utah/A00461135/2009(H7N1) A/northern shoveler/Mississippi/A00630207/2009(H7N6) A/chicken/Jiangxi/JXA132304/2013(H9N2)a A/mallard/South Dakota/A00536114/2007(H10N7) A/mallard/New York/A00919062/2009(H10N7) A/chicken/Jiangxi/JXA132717/2013 (H10N8)a Avian Newcastle disease virus (strain 2013 pt#3-3) Infectious bursal disease virus (strain P13-03308) Infectious bronchitis virus (strain P14-00957ct) Reovirus (strain P14-00932) Note:a: The RNA from these viruses was extracted directly from a clinical swab sample; for all others viruses, the RNA was extracted from virus isolates. Table S1. Viruses used to determine the specificity of a Eurasian-lineage H10 specific quantitative RT-PCR method

 DownLoad:
DownLoad: