











-
Potato (Solanum tuberosum L.) is the third most important food crop belonging to the family Solanaceae (Masum et al., 2011; Bae et al., 2012; Barrell et al., 2013), which is cultivated all over the world (Czajkowski et al., 2011). Over 1 billion people have potato as their staple diet (Barrell et al., 2013). Bacterial wilt is caused by the soil-born Gram-negative bacterium Ralstonia solanacearum (Fujiwara et al., 2011) (synonym Pseudomonas solanacearum) and is the second most serious bacterial disease in potato. The crop yield may decline 80% to 100% after infection by the pathogen. It is therefore important to develop effective ways for controlling potato bacterial wilt.
R. solanacearum cells can survive for many years in association with alternate hosts and are easily disseminated via soil, contaminated irrigation water, surface water, farm equipment, and infected biological material. Soil fumigation with methyl bromide, vapam, or chloropicrin is normally used as a treatment, but with limited efficacy. Bactericides, such as antibiotics and copper-based compounds (Jones et al., 2012), have been commonly used to control the disease. However, resistance of R. solanacearum to these antimicrobial compounds has been reported (Hayward, 1991; Stratton et al., 2003; Wicker et al., 2007). Crop rotation has also been applied to control bacterial diseases. Unfortunately, the diverse and broad host range of R. solanacearum, and limited land area available, make it impossible to control the disease through crop rotation (Denny et al., 2007; Sagar et al., 2013; Farag et al., 2015). Pretreatments of plants with avirulent bacteriocin-producing strains and microbial antagonism to control bacterial wilt have also been investigated (Chen et al., 1984; Ciampi-Panno et al., 1989), but complete control of the disease has yet to be achieved. Due to the limited effectiveness of the current integrated management strategies, bacterial wilt continues to be an economically serious problem for potato production in many tropical and subtropical regions of the world (Wicker et al., 2007; Sagar et al., 2013).
Recently, lytic bacteriophages have been found to be effective and safe for controlling bacterial diseases (Fujiwara et al., 2011; Addy et al., 2012). Addy et al. (2012) used a filamentous phage, ØRM3, to control tomato bacterial wilt caused by R. solanacearum under greenhouse conditions. Notable advantages of using bacteriophages include safety to animals and the environment, as well as ease of production. Bacteriophages are also self-replicating and self-limiting, and are able to overcome bacterial resistance (Carlton, 1999; Loc-Carrillo et al., 2011). However, bacteria in many cases are genetically diverse and, therefore, differ in susceptibility to phages. At the same time, bacteria can easily mutate to become resistant to a particular phage (Ye et al., 2010). Therefore, a cocktail of lytic bacteriophages isolated from diverse environments may have the potential for application as a universal biocontrol agent with reduced likelihood for the emergence of resistance. As far as we know, no studies have been reported on developing bacteriophage cocktails for the biocontrol of potato bacterial wilt.
In this work, twelve lytic bacteriophages were tested against R. solanacearum strains isolated from different water sources. A bacteriophage cocktail was formulated and tested for efficacy in the decontamination of pathogen-contaminated sterilized soil and protection of potato plants growing in garden pots.
-
One reference strain, G (GIM1.74, bought from Guangdong Microbiology Culture Center, China), and three field isolates of R. solanacearum (PS-X4-1, PS-X10-2, and PS-X13-1) from wilted potato plants (Yunnan province, China) were used in this study. The reference strain G was used in the isolation of phages from water samples and in the production of the bacteriophages. The field isolate X (PS-X4-1) was used in the pot tests. For routine experiments, bacteria were grown at 28 °C in CPG broth (0.1% casamino acids, 1% tryptone, 0.5% glucose, pH 7.0) or on CPG agar plates (CPG broth with 1.5% agar) for 24–48 h (Fujie et al., 2010). The strains were kept in 20% glycerol at –80 °C for long-term storage.
-
Bacteriophages were isolated from various surface-water samples collected in China and Kenya, following previously described procedures (Hanumanthappa et al., 2013). The presence of phages in the samples was analyzed by the soft agar overlay protocol (Adams et al., 1959). Briefly, a water sample was filtered through a membrane filter (0.22 μm pore size; Tianjing Jingteng Co. Ltd., China). Then, 1 mL of an equivalent-volume mixture of the bacteria (1×108 CFU/mL) and the filtered water solution was mixed with 4 mL of 0.7% soft agar. Finally, the mixture was overlaid onto CPG plates containing 1.5% agar (Murugaiyan et al., 2011). The plates were incubated at 28 °C for 12 h before plaque detection. Single-plaque isolates were chosen and purified at least 5 times by spreading the mixtures of the single phage and the host bacteria onto CPG agar plates. Only those phages that formed stable plaques during each round were deemed as lytic phages and selected for further analysis.
To amplify the phage, an overnight culture of bacterial cells grown in CPG medium was diluted 100-fold with 100 mL fresh CPG medium in a 500 mL flask. When the cultures reached an OD600 (the optical density of the bacteria solutions at 600 nm) reading of 0.5, a bacteriophage was added at a multiplicity of infection (MOI) of 0.001–1.0 (usually 0.01–0.1). After culturing for a further 12 h, the cells were removed by centrifugation at 8000 × g for 15 min at 4 °C. The supernatant was passed through a 0.22 μm membrane filter, and the phage solutions were stored in 20% glycerol at 4 °C and –80 °C until used.
-
The MOI is the ratio between phage particles and potential bacterial host cells (Ji et al., 2015). R. solanacearum host cells were infected with phage isolates at ratios ranging from 0.00001 to 0.1, increasing 10 folds gradually. The phage solution and host broth were blended together with 4 mL 0.7% water–agar, and then the mixtures were poured onto CPG solid plates, the plates were incubated inverted at 28 °C for 12–24 h, and single plaques counted.
-
The four different bacterial strain isolates (R. solanacearum GIM1.74, PS-X4-1, PS-X10-2, and PS-13-1) under test were cultured overnight (cell density, 1010 CFU/mL) in CPG liquid medium. Then, 500 μL broth culture of the bacterial strains was mixed with 4 mL of 0.7% soft agar and overlaid onto CPG plates containing 1.5% agar to prepare the R. solanacearum lawns. Ten microliters of phage suspensions (1010 CFU/mL) were spotted onto the R. solanacearum lawns, which were then incubated at 28 °C for 12 h (Popova et al., 2012; Kwiatek et al., 2012). Bacterial sensitivity to different phages was defined by the clarity of the positions where the phage solutions were deposited.
-
The adsorption tests and one-step growth experiments were undertaken as described by Dr. Czajkowski et al. (2014) and Dr. Simoliunas et al. (2013). Adsorption of the phages was measured as follows: infection of R. solanacearum GIM1.74 of mid-exponential phase in CPG medium at optimal MOIs of different phages, followed by gentle mixing, and 100 μL of the mixtures was taken instantly to determine the initial phage titers. Incubation lasted 30 min, and 100-μL volumes of the samples were taken at intervals of 3 min and diluted immediately into 900 μL of cooled phage buffer. The diluted samples were centrifuged at 12,000 × g for 5 min at 4 °C. The titers of unadsorbed phages in the supernatant were determined after serial dilution. The one-step growth curve was carried out using a previously described protocol (Hsieh et al., 2011).
-
The morphology of three selected phages was analyzed by transmission electron microscopy (TEM). Approximately 1×109 PFU/mL phages purified by CsCl density-gradient centrifugation (SW41 rotor, Beckman LE-80K, USA) were dropped onto 200 mesh copper grids and stained with 2% (w/v) uranyl acetate. After drying, morphology analysis was performed using a Hitachi H-7000FA electron microscope (Japan).
-
The effects of the phages on bacterial growth were measured by monitoring the OD600 for 12 h post-infection with each phage. Inocula (250 μL) of R. solanacearum G cells of mid-exponential phase (1010 CFU/mL) were mixed with 250 μL of solutions of the twelve lytic phages alone or in combination at a MOI of 1, then 500 μL of CPG liquid medium was added to the mixture (Fujiwara et al., 2011). The OD600 of the final mixtures was monitored continuously with a micro-plate reader (SynergyH1, BioTek, USA) at 28 °C, over 12 h.
Based on the lytic curves of the single bacteriophages, we divided the 12 bacteriophages into four types according to the trends of the curves. We selected different numbers of bacteriophages from every type and combined them to measure the lysis kinetics of the bacteriophage cocktails (Supplementary Table S1), as described above.
-
To make the phage cocktails, the selected lytic bacteriophages were propagated in CPG medium co-cultured with host broth, and resuspended in phage buffer at a density of 108 PFU/mL. Then, each phage was mixed at an equivalent volume to produce phage cocktails. Among them, the best mixture, containing equivalent volumes of six bacteriophages (P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-7, P-PSG-8, P-PSG-9), was named the P1 phage cocktail.
A sensitive potato (S. tuberosum L.) type, Favorita (obtained from Zhongjia Agricultural Biotechnology Co., Ltd, Inner Mongolia, China), was selected for the pathogen inoculation and bacteriophage control experiments. Test-tube grown plantlets were transplanted into pots (mouth diameter, 33 cm; root diameter, 25 cm; height, 25 cm) containing solid growth medium (vermiculite, manure, humectant, pH 5.0–6.0) under net-shed conditions. The temperature outside was 20 ± 5 °C, and the relative humidity ranged from 50 % to 80%.
The experiments were divided into three general categories: control, prevention, and therapeutic. Under each category, different test groups were set up with five plants in each group. Inoculation of pathogens or phage formulations was carried out similarly to methods described previously (Fujie et al., 2010). The solutions were injected with a sterile 1-mL syringe (HengKang Medical Apparatus and Instruments Co., Ltd, Wuhan, China) into the second internode on the major stem of 4-week-old potato plants (with 4–6 leaves).
In the control category, the impacts of P1 and two bacterial strains (G and X) on the plants were tested by injection of P1 only (1×108 PFU/mL each) or each of the two R. solanacearum strains (G and X) (2×108 CFU/mL) separately to the plants. Sterile water (SW) was used as the control sample. The four groups in this first category were separately named as P1, G, X, and SW.
In the prevention category, the prevention efficiencies of P1 were tested by inoculating the plants first with P1 (1×108 PFU/mL each), and then with R. solanacearum G and X (2×108 CFU/mL), 2 h after the injection of P1. The groups were named as P1-G and P1-X, respectively.
In the therapeutic category, the therapeutic effectiveness of P1 was tested by inoculating the plants first with R. solanacearum G and X (2×108 CFU/mL), and then inoculating the plants 3, 6, 9, 12, and 15 days later with P1 (1×108 PFU/mL each), for a total of 5 times. The inoculated plants were coded and observed daily for wilt progress under the different treatments (Roberts et al., 1988).
-
R. solanacearum G grown in CPG liquid medium was harvested at the exponential growth phase by centrifugation (8000 × g for 15 min at 4 °C), and the cells were washed and resuspended in distilled water (Bhunchoth et al., 2015). A 5-mL inoculum of the cell suspension (6.0×108 CFU/mL) was added to autoclaved soil (30 g) in a 50-mL flask and mixed well. At the same time, 5 mL of SW was added into another flask with 30 g of autoclaved soil as the control. One day later, 5 mL of P1 (6.0×109 PFU/mL each) was sprayed on the surface of the contaminated soil at a MOI of 10 (approx. 1.0×109 PFU/g soil). A similar amount of P1 was sprayed onto the control soil treated with water. The soil mixtures were kept at room temperature and small portions (1 g) of the samples were collected on different days to monitor the number of GIM1.74 cells in the soils. The bacterial cell number was counted on CPG agar plates using the spread-plating method.
-
About 60 water samples were collected from geographically different water sources in Kenya and China during the years 2014 and 2015. These samples were assayed for the presence of lytic bacteriophages against R. solanacearum strain G as the host. Approximately 30% of the tested water samples (18 samples) produced lytic plaques on assay plates. Finally, we obtained 12 lytic phages that were virulent to all three field isolates. The lytic phages had different characteristics, as summarized in Table 1. In addition to R. solanacearum strain G, all the phages were found to be able to lyse the other three bacterial isolates (PS-X4-1, PS-X10-2, and PS-13-1), showing that they might have relatively wide spectra against R. solanacearum. Adsorption kinetics curves and one-step growth curves of all the 12 phages are presented in Supplementary Figure S1 and Supplementary Figure S2, respectively. As presented in Figure 1, TEM analysis showed that two of the selected phages (P-PSG-1 and P-PSG-7) belong to family Siphoviridae, while P-PSG-6 did not have a tail and spikes; therefore, it belongs to family Cystoviridae.
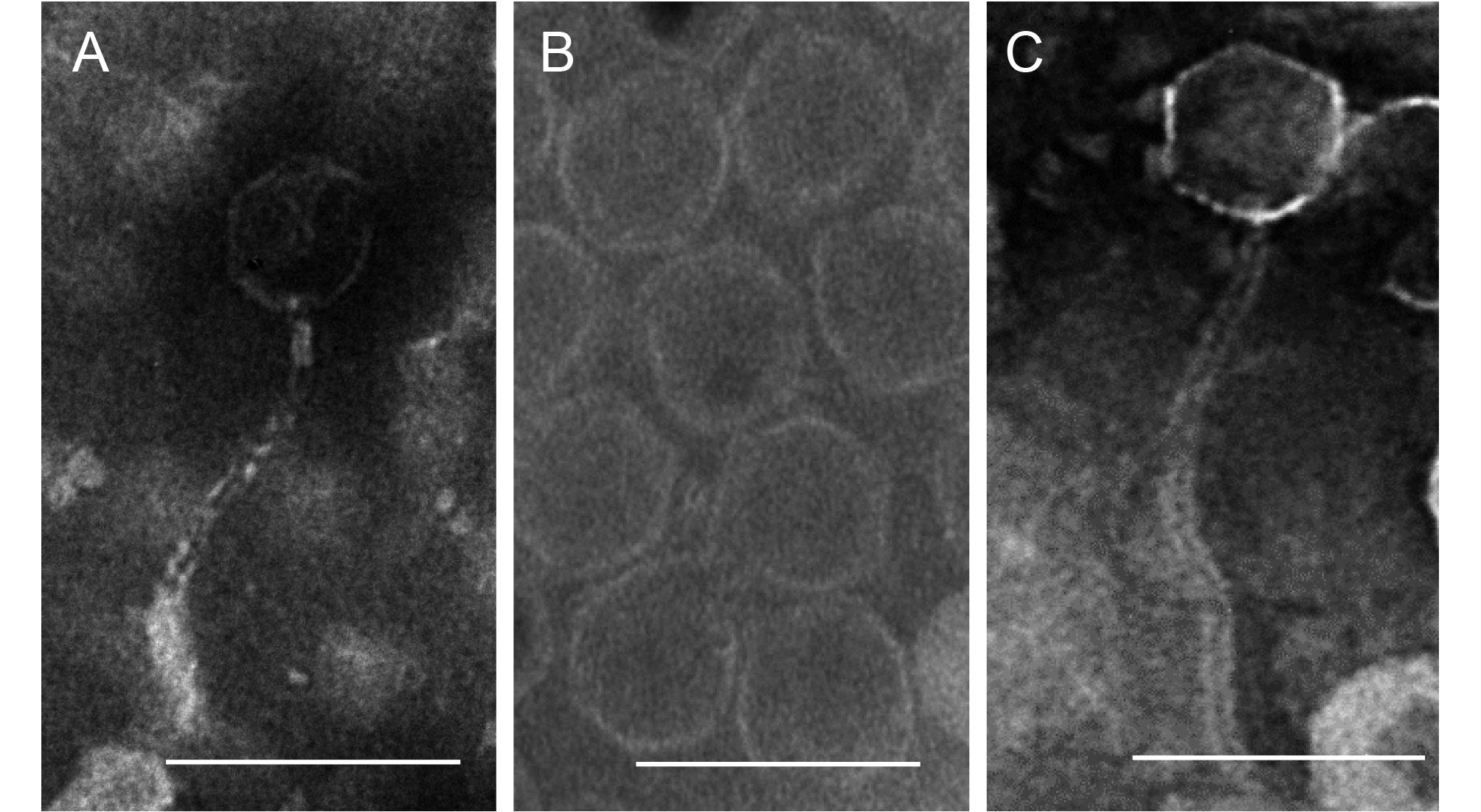
Figure 1. TEM images of the phages P-PSG-1 (A), P-PSG-6 (B), and P-PSG-7 (C). Bar size: 100 nm.
Phage name Isolate source Diameter#
(cm)Optimal MOI Latent period*
(min)Burst size*
(ratio)P-PSG-1 Zhengdian, Hubei
Province, China0.4 10–3 / 10.5 P-PSG-2 Zhengdian, Hubei
Province, China0.6 10–2 / 15.5 P-PSG-3 East-Lake, Hubei
Province, China0.5 10–3 20 1200 P-PSG-4 East-Lake, Hubei
Province, China0.5 10–3 20–30 65.8 P-PSG-5 Qingshan Lake, Hubei
Province, China0.5 10–4 / 2.7 P-PSG-6 BuHe, Anhui
Province, China0.1 10–4 15 191.4 P-PSG-7 River Sare,
Migori, Kenya0.5 10–5 90 200 P-PSG-8 River Migori,
Migori, Kenya0.5 10–5 50–70 53.0 P-PSG-9 Stella Well,
Kisumu, Kenya0.4 10–3 70 330 P-PSG-10 River Rianyago,
Migori, Kenya0.4 10–4 70 633.3 P-PSG-11 River Nyambira,
Migori, Kenya0.4 10–4 80 124.8 P-PSG-12 Mwamogesha Spring,
Migori, Kenya0.4 10–5 10–20 198.6 Note: #, diameter of plaque; *, latent period and burst size are derived from adsorption kinetics curve and one-step growth curves. Table 1. Partial characterization of the 12 bacteriophages
-
The 12 phages could be roughly classified into four types based on the curves shown in Figure 2. P-PSG-3 and P-PSG-4 belong to the first group. These two phages inhibited growth of the host bacteria immediately after infection. The OD600 decreased to baseline within 2.5 h post-infection (Figure 2A). The second group consists of six phages (P-PSG-1, P-PSG-8 to P-PSG-12). The OD600 increased to the maximum within the first 1 h, then remained stable for about 1 h, and finally decreased to baseline within 6 h post-infection by these phages (Figure 2B). The third group comprises P-PSG-2 and P-PSG-7. The OD600 increased to the maximum within the first 3h, and then gradually decreased to baseline more than 12 h post-infection (Figure 2C). The last group consists of P-PSG-5 and P-PSG-6. The OD600 increased to the maximum within the first 5 h, and then gradually decreased, following the same trend as the group comprising P-PSG-2 and P-PSG-7 (Figure 2D). The OD600 of the bacterial control uninfected with any bacteriophages increased gradually within the first 8 h to the maximum (about 0.65) and was still stable after 12 h when the experiment was terminated (Figure 2F).
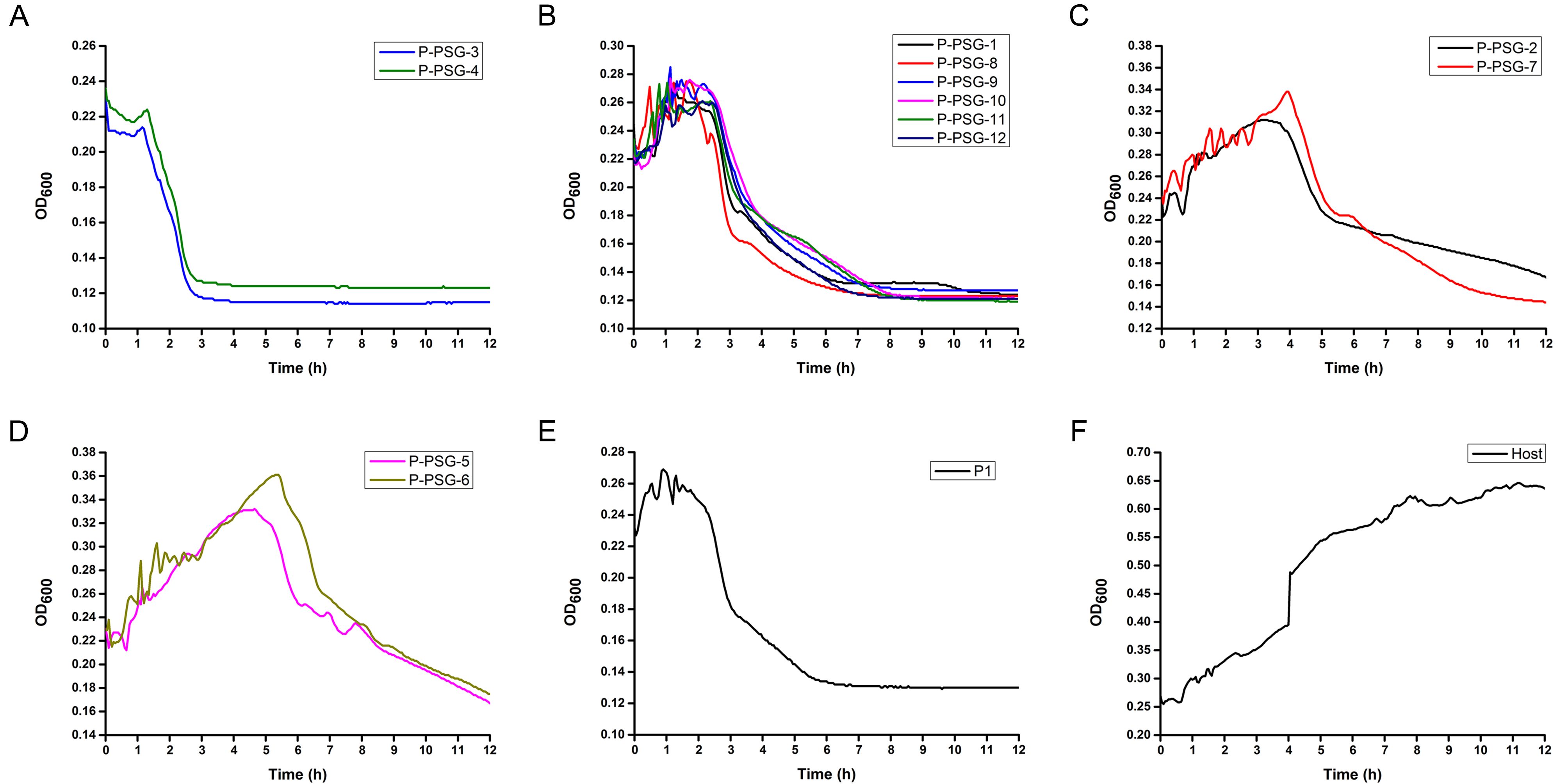
Figure 2. Growth curves of R. solanacearum strain G uninfected (F), or after infection with different bacteriophages (A–D), or the phage cocktail P1 (E). Inocula (250 μL) of R. solanacearum G cells of mid-exponential phase (1010 CFU/mL) were mixed with 250 μL of solutions of the 12 lytic phages alone or in combination at a MOI of 1, then 500 μL of CPG liquid medium was added into the mixture. The absorption of the final mixtures at 600 nm (OD600) was monitored continuously with a micro-plate reader at 28 °C, over 12 h.
-
Based on the above lytic kinetics, bacteriophages from each of the groups were selected for mixing to produce cocktails (Supplementary Table S1). Generally, all the cocktails showed better inhibition of the growth of the host bacterial strain G than single bacteriophages. The initial phase of increasing OD600 was reduced to less than 1 h post-infection by the cocktails. The OD600 value decreased rapidly to the baseline within 4 h. Exceptionally, of all the cocktails tested, one phage cocktail, P1, consisting of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-7, P-PSG-8, and P-PSG-9, left the lowest number of live bacterial cells at 12 h post-infection (Figure 2E). Therefore, the phage cocktail P1 was chosen for the following experiments to test for efficacy in the prevention and treatment of potato wilt, as well as in the decontamination of pathogen-contaminated soil.
-
Generally, two approaches could be used to prevent bacterial wilt. One is to pretreat the potato plants with the cocktail, so that the plants could be prevented from later infections by the bacteria pathogen. Another is to decontaminate pathogen-contaminated soil before planting potato plants.
We first tested whether pretreating the potato plants with the P1 phage cocktail could prevent bacterial wilt development by injecting R. solanacearum into the stem of the potato plants. Plants injected with P1 (P1 group of bacteriophages) or SW (SW group) grew normally and did not develop any symptoms of bacterial wilt. The results show that P1 is quite safe to the plants. In contrast, as shown in Table 2, all the plants developed wilt after inoculation with either R. solanacearum strain G (G group) or strain X (X group). The times to the first observation of wilt symptoms were different for different plants. Generally, the plants developed wilt within 3–11 days after inoculation of R. solanacearum G, while the times ranged from 3 to 10 days after inoculation of R. solanacearum X. However, once they showed wilt symptoms, the plants were completely dead within 3–5 days when infected with R. solanacearum G, and within 2–3 days when infected with R. solanacearum X. These results show that the virulence of R. solanacearum X is stronger than that of R. solanacearum G.
Group Plant no. Day of first
observation of wilt
after inoculationDay of complete
wilt after
inoculationGroup Plant no. Day of first
observation of wilt
after inoculationDay of complete
wilt after
inoculationG 1 5 10 X 1 4 6 2 5 9 2 5 7 3 7 11 3 6 9 4 10 15 4 7 10 5 11 16 5 10 12 P1-G 1 / / P1-X 1 / / 2 / / 2 5 10 3 / / 3 / / 4 / / 4 / / 5 10 18 5 / / G-P1-3d 1 / / X-P1-3d 1 7 13 2 / / 2 7 14 3 / / 3 5 10 4 / / 4 3 6 5 3 8 5 9 16 G-P1-6d 1 / / X-P1-6d 1 7 12 2 6 / 2 3 5 3 / / 3 10 16 4 7 / 4 5 9 5 / / 5 3 6 G-P1-9d 1 5 9 X-P1-9d 1 6 9 2 7 12 2 3 6 3 / / 3 3 6 4 9 18 4 7 9 5 / / 5 5 7 G-P1-12d 1 5 9 X-P1-12d 1 3 5 2 6 10 2 4 6 3 4 9 3 5 7 4 10 15 4 3 6 5 / / 5 9 12 G-P1-15d 1 5 9 X-P1-15d 1 5 7 2 9 14 2 9 11 3 4 8 3 3 5 4 4 9 4 7 10 5 9 14 5 3 6 Table 2. Disease progression in potato plants following inoculation of R. solanacearum under different bacteriophage treatments
Injecting P1 into the potato plant stems 2 h before inoculation with either R. solanacearum G (P1-G group) or X (P1-X group) was found to prevent the development of wilt. As shown in Table 2, the morbidities of the plants (both P1-X group and P1-G group) were reduced to 20%. These results showed that the bacteriophage cocktail P1 could prevent infection by R. solanacearum in potato plants.
P1 also showed a capability to reduce the density of live bacteria in pathogen-contaminated sterilized soils. The CFU of the bacteria in the contaminated soils decreased more than 5-fold after spraying P1 into the soils, compared with the control (Figure 3). One week post-spray, the number of live bacteria was only about 2% of those in the control soils.
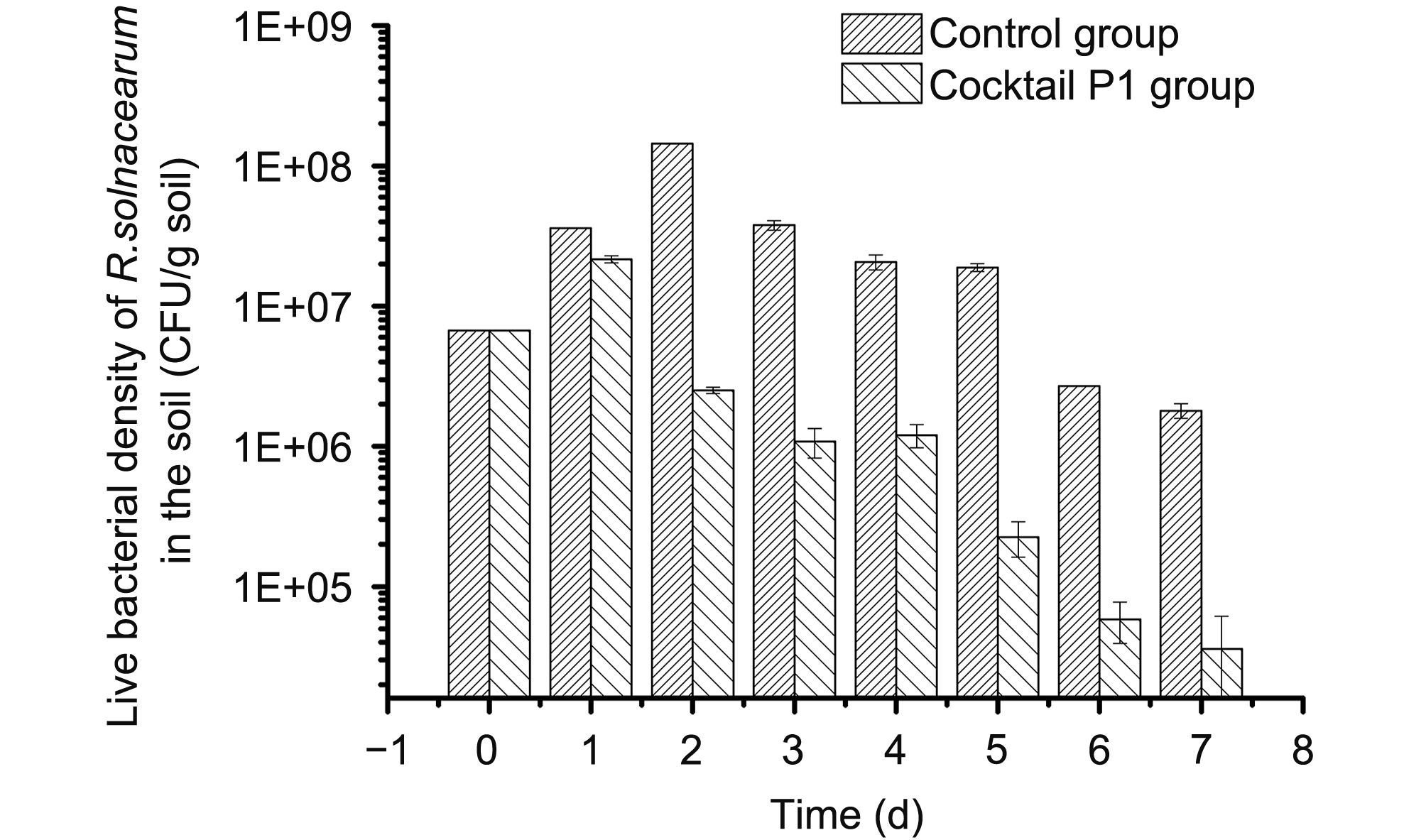
Figure 3. Bioassay for the bactericidal effect of the P1 cocktail: decontamination of pathogen-contaminated soil by P1. R. solanacearum G grown in CPG liquid medium was harvested at exponential growth phase as described in the Methods. A 5-mL inoculum of cell suspension (6.0×108 CFU/mL) or SW (as a control) was added to autoclaved soil (30 g) in a 50-mL flask and mixed well. One day later, 5 mL of P1 (6.0×109 PFU/mL each) was sprayed on the surface of the control soil and the contaminated soil at a MOI of 10 (approx. 1.0×109 PFU/g soil). The soil mixtures were kept at room temperature and small portions (1 g) were collected on different days to monitor the number of GIM1.74 cells in the soils. Bacterial cell numbers were counted on CPG agar plates using the spread-plating method.
-
We further tested whether P1 could be used to treat bacterial wilt after the plants were inoculated with the pathogenic bacteria. As shown in Table 2, after inoculation of R. solanacearum G, the wilt could be controlled by early injection of P1 at 3 and 6 days post-inoculation, except for one plant in the group G-P1-3d, which developed the first sign of wilt on the same day as P1 was applied. For the more virulent strain X, all the infected plants died due to the wilt. However, earlier injection of P1 could lengthen the dying time. For example, in the group X-P1-3d, the times from the first signs of wilt to complete wilt were delayed to 4–5 days, instead of 2–3 days without the treatment.
-
As bacteriophages are limited to particular strains within a single bacterial species, it is generally considered that a correct phage cocktail would be needed to achieve effective disease control. In the current study, 12 lytic phages from different regions in Kenya and China were isolated. We hoped that the diversity of phages would provide a good basis for formulating a phage cocktail that would lyse most R. solanacearum strains to prevent potato bacterial wilt. From the curves of the lytic dynamics shown in Figure 2, these phages showed different inhibition patterns against growth of the bacteria, which might reflect their differences when interacting with the host bacteria. Interestingly, the curve patterns seemed to show certain connections to the water sources from which they were isolated. For example, for the six phages in the second group, five of them (P-PSG-8, P-PSG-9, P-PSG-10, P-PSG-11, P-PSG-12) were isolated from the waters sampled in Kenya. Furthermore, these phages were isolated from water sources, not from soils as in previous studies (Fujiwara et al., 2011). The fact that lytic bacteriophages could be isolated from approximately 30% of the tested water samples (18 samples) indicates that surface waters are a rich resource for lytic bacteriophages against R. solanacearum.
Currently, there are many guidelines for selecting a phage to be used in a phage cocktail, but the most important requirement is to make sure that the phage cocktail can kill the target bacteria rapidly and effectively. However, so far there are few studies that have been reported on how to formulate and assess the efficacy of a phage cocktail. We found that using lytic dynamics curves could be a good way to achieve this. First, the lytic dynamics curve could reflect the effects of an individual phage on the growth of the target bacteria. Second, based on the lytic dynamics curves, one could choose phages with different patterns to formulate a phage cocktail comprising a diversity of bacteriophages, which might work on most of the target bacterial strains. Finally, the lytic dynamics could be used to assess the effect of the phage cocktail on growth of the bacteria. As shown in Figure 2, a more effective inhibition of the growth of strain GIM1.74 by the phage cocktail P1 than by any individual bacteriophage could be clearly seen. The second criterion for choosing phages to formulate a cocktail would be to include phages acting on a wide range of R. solanacearum strains. Due to the limited number of R. solanacearum field isolates available in our laboratory, the lytic spectrum of the phage cocktail P1 needs further testing.
Preliminary pot tests showed that using the phage cocktail P1 is feasible to prevent potato bacterial wilt either by injecting phages into the plants or by decontamination of the soil. The presence of the phages could reduce the infection rates or greatly limit the number of bacteria in the soil. However, the treatment efficacies of P1 depended upon the timing of application of the phages, and the susceptibility of the plants to the bacterial wilt, as well as upon the virulence of the bacteria infected. The earlier the phages were applied, the better the outcome of the treatment was (Table 2). At the same time, it seems that bacteriophages could only delay or stop the dying of the plants due to bacterial wilt, but could not repair the damage caused by the wilt. Therefore, if the virulence of the infecting bacteria is strong (such as in the case of strain P-X4-1 in this study), the plants could be damaged rapidly. Application of the phages could only delay death (from 7 days without phages to 10 days with the phage therapy in the X-P1-3d group of this study). Therefore, these results show that it is better to use the phages as a prevention agent; or, in plants already infected, it is important to apply the phage therapy as soon as possible, preferably bacterial wilt is observed. Given that there are many stages involved in potato cultivation, it may be most convenient to apply the phages by immersing potato tubers into a phage solution before planting in the soil, or by using phage solutions to water the potato plants. Further studies are in progress to find suitable and economic ways for applying phages to prevent potato bacterial wilt.
In conclusion, 12 lytic bacteriophages were isolated from different surface waters in Kenya and China in order to study their potential to control potato bacterial wilt. Based on the lytic dynamics curves, a phage cocktail P1 was formulated which showed enhanced speed and efficacy in the lysis of R. solanacearum bacteria pathogens. The preliminary pot tests showed that the phage cocktail was very effective in preventing potato bacterial wilt. However, further studies are needed to develop simple and economic methods of applying the phage cocktail in the field.
-
This study was supported financially by the Sino-Africa Joint Research Center (SAJC201605) and the Chinese Academy of Sciences (ZDRW-ZS-2016-4). We thank Dr. Ding Gao from the core facility center, Wuhan Institute of Virology, CAS for the help on TEM observation.
-
The authors declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by any of the authors.
-
CHW, RFZ, and HPW designed the experiments. CHW, JLL, ANM, JPY, CHY carried out the experiments. CHW, RFZ, and HPW analyzed the data. CHW, FBM and HPW wrote the paper. All authors read and approved the final manuscript.
Supplementary Figures and Table are available on the websites of Virologica Sinica: www.virosin.org; link.springer.com/journal/12250.
-
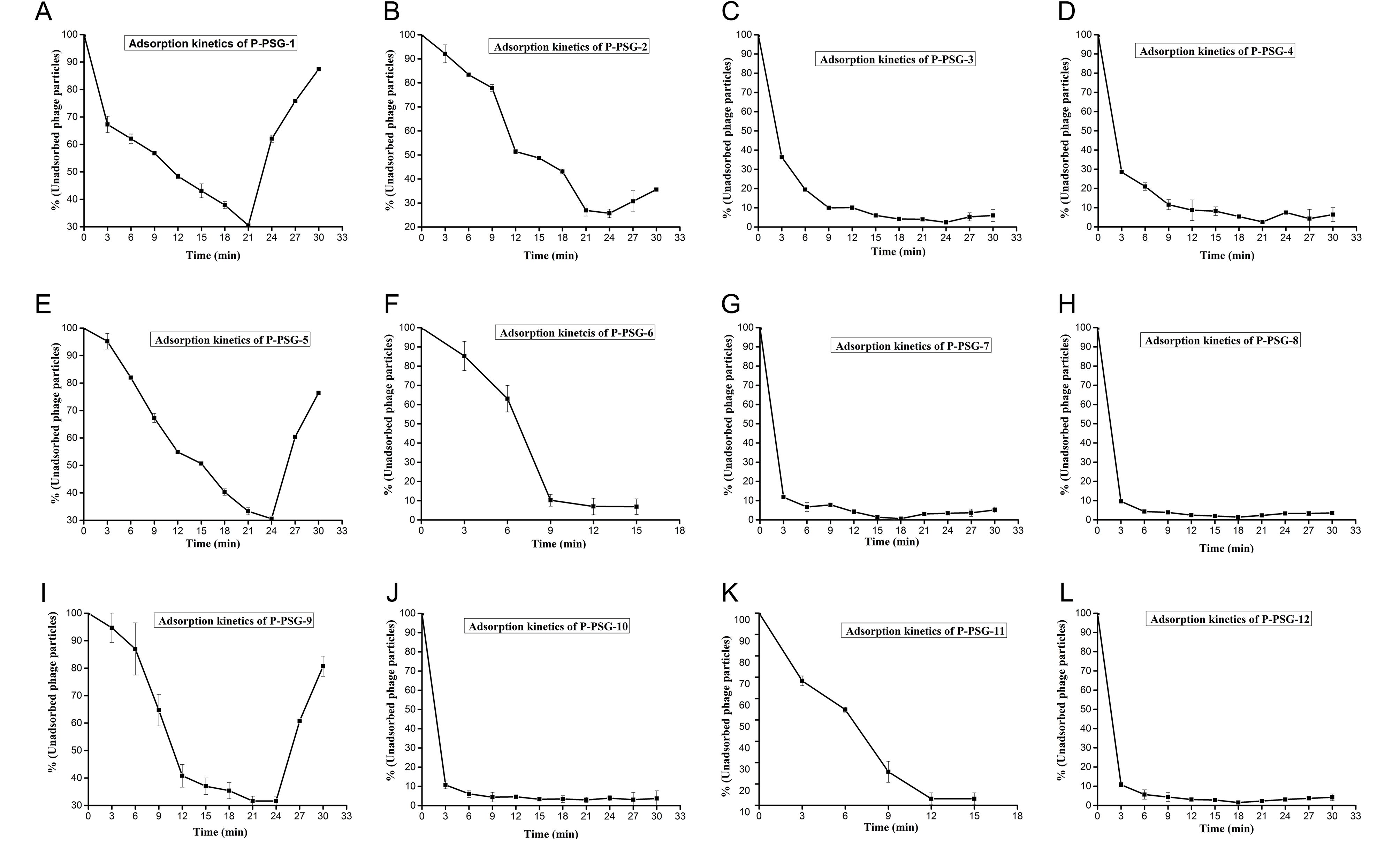
Figure S1. Adsorption kinetics curves for the 12 bacteriophage isolates.

Figure S2. One-step growth curves for the 12 bacteriophage isolates.
Group name Titer (PFU) MOI Source P-PSG-1 3.06×1010 1 Single P-PSG-1 P-PSG-2 1.84×1010 1 Single P-PSG-2 P-PSG-3 8.0×1010 1 Single P-PSG-3 P-PSG-4 9.8×1010 1 Single P-PSG-4 P-PSG-5 7.2×1010 1 Single P-PSG-5 P-PSG-6 3.0×1010 1 Single P-PSG-6 P-PSG-7 1.0×1010 1 Single P-PSG-7 P-PSG-8 1.8×1010 1 Single P-PSG-8 P-PSG-9 1.3×1010 1 Single P-PSG-9 P-PSG-10 1.42×1010 1 Single P-PSG-10 P-PSG-11 1.4×1010 1 Single P-PSG-11 P-PSG-12 2.42×1010 1 Single P-PSG-12 P-PSG-(1/2) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2 P-PSG-(1/8) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-8 P-PSG-(3/10) 1010 1 Equivalent volume mixture of P-PSG-3, P-PSG-10 P-PSG-(6/11) 1010 1 Equivalent volume mixture of P-PSG-6, P-PSG-11 P-PSG-(1/4/7) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-4, P-PSG-7 P-PSG-(1/7/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-7, P-PSG-12 P-PSG-(2/9/10) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-9, P-PSG-10 P-PSG-(3/5/12) 1010 1 Equivalent volume mixture of P-PSG-3, P-PSG-5, P-PSG-12 P-PSG-(4/8/9) 1010 1 Equivalent volume mixture of P-PSG-4, P-PSG-8, P-PSG-9 P-PSG-(5/7/9) 1010 1 Equivalent volume mixture of P-PSG-5, P-PSG-7, P-PSG-9 P-PSG-(6/8/11) 1010 1 Equivalent volume mixture of P-PSG-6, P-PSG-8, P-PSG-11 P-PSG-(1/2/7/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-7, P-PSG-12 P-PSG-(1/3/8/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-3, P-PSG-8 P-PSG-12 P-PSG-(2/3/4/5) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5 P-PSG-(2/4/9/11) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-4, P-PSG-9, P-PSG-11 P-PSG-(3/5/10/12) 1010 1 Equivalent volume mixture of P-PSG-3, P-PSG-5, P-PSG-10, P-PSG-12 P-PSG-(6/8/10/12) 1010 1 Equivalent volume mixture of P-PSG-6, P-PSG-8, P-PSG-10, P-PSG-12 P-PSG-(1/2/3/4/6) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-6 P-PSG-(1/3/6/8/11) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-3, P-PSG-6, P-PSG-8, P-PSG-11 P-PSG-(2/3/4/5/8) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-8 P-PSG-(2/3/4/6/11) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-6, P-PSG-11 P-PSG-(2/4/5/7/9) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-4, P-PSG-5, P-PSG-7, P-PSG-9 P-PSG-(3/6/8/10/12) 1010 1 Equivalent volume mixture of P-PSG-3, P-PSG-6, P-PSG-8, P-PSG-10, P-PSG-12 P-PSG-(4/6/7/9/11) 1010 1 Equivalent volume mixture of P-PSG-4, P-PSG-6, P-PSG-7, P-PSG-9, P-PSG-11 P-PSG-(1/2/3/4/5/6) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-6 P-PSG-(1/2/3/7/8/9)=P1 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-7, P-PSG-8, P-PSG-9 P-PSG-(2/3/6/7/9/11) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-3, P-PSG-6, P-PSG-7, P-PSG-9, P-PSG-11 P-PSG-(3/4/5/8/9/10) 1010 1 Equivalent volume mixture of P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-8, P-PSG-9, P-PSG-10 P-PSG-(4/5/6/10/11/12) 1010 1 Equivalent volume mixture of P-PSG-4, P-PSG-5, P-PSG-6, P-PSG-10, P-PSG-11, P-PSG12 P-PSG-(7/8/9/10/11/12) 1010 1 Equivalent volume mixture of P-PSG-7, P-PSG-8, P-PSG-9, P-PSG-10, P-PSG-11, P-PSG-12 P-PSG-(1/2/3/4/6/7/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-6, P-PSG-7, P-PSG-12 P-PSG-(1/2/3/4/6/11/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-6, P-PSG-11, P-PSG-12 P-PSG-(2/3/4/5/8/9/10) 1010 1 Equivalent volume mixture of P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-8, P-PSG-9, P-PSG-10 P-PSG-(1/2/3/4/5/6/7/8/9/10/11/12) 1010 1 Equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-6, P-PSG-7P-PSG-8, P-PSG-9, P-PSG-10, P-PSG-11, P-PSG-12 Host broth only 1010 (CFU) / No bacteriophages present Bacteriophage cocktail only 1010 / No host; equivalent volume mixture of P-PSG-1, P-PSG-2, P-PSG-3, P-PSG-4, P-PSG-5, P-PSG-6, P-PSG-7P-PSG-8, P-PSG-9, P-PSG-10, P-PSG-11, P-PSG-12 Table S1. Bacteriophage cocktails used in lysis kinetics analyses
Developing a bacteriophage cocktail for biocontrol of potato bacterial wilt
- Cuihua Wei 1,2 ,
- Junli Liu 3 ,
- Alice Nyambura Maina 4,5 ,
- Francis B Mwaura 4 ,
- Junping Yu 1 ,
- Chenghui Yan 1,2 ,
-
Ruofang Zhang
3,,
 ,
, -
Hongping Wei
1,6,,
- Received Date: 28 March 2017
- Accepted Date: 18 October 2017
- Published Date: 16 November 2017
Abstract: Bacterial wilt is a devastating disease of potato and can cause an 80% production loss. To control wilt using bacteriophage therapy, we isolated and characterized twelve lytic bacteriophages from different water sources in Kenya and China. Based on the lytic curves of the phages with the pathogen Ralstonia solanacearum, one optimal bacteriophage cocktail, P1, containing six phage isolations was formulated and used for studying wilt prevention and treatment efficiency in potato plants growing in pots. The preliminary tests showed that the phage cocktail was very effective in preventing potato bacterial wilt by injection of the phages into the plants or decontamination of sterilized soil spiked with R. solanacearum. Eighty percent of potato plants could be protected from the bacterial wilt (caused by R. solanacearum reference strain GIM1.74 and field isolates), and the P1 cocktail could kill 98% of live bacteria spiked in the sterilized soil at one week after spraying. However, the treatment efficiencies of P1 depended on the timing of application of the phages, the susceptibility of the plants to the bacterial wilt, as well as the virulence of the bacteria infected, suggesting that it is important to apply the phage therapy as soon as possible once there are early signs of the bacterial wilt. These results provide the basis for the development of bacteriophage-based biocontrol of potato bacterial wilt as an alternative to the use of antibiotics.

 DownLoad:
DownLoad: