












HTML
-
In early 2013, the first case of a human infection with avian influenza A (H7N9) virus was reported in the Yangtze River Delta region of China (Liu et al. 2013). Since early 2013, a total of 1533 laboratory-confirmed human infections with avian H7N9 viruses have been reported to the World Health Organization (http://www.who.int/csr/don/28-june-2017-ah7n9-china/en/), with five epidemic waves of this virus occurring in China. Human infections with the highly pathogenic H7N9 influenza virus have emerged particularly in China (Zhang et al. 2017). The increasing numbers of human infections with H7N9 influenza viruses raises concerns that these viruses will continue to be a potential pandemic threat to public health.
It has been shown that the H7N9 virus responsible for the 2013 Chinese outbreak originated from gene reassortment among H7, N9 and H9N2 avian influenza viruses (AIVs), and it appears that the internal genes of the H7N9 viruses are most closely related to a wild land bird virus (A/brambling/Beijing/16/2012, H 9N2) (Chen et al. 2013; Liu et al. 2015). Therefore, wild land birds may have played a role in generating the 2013 H7N9 viruses. During the ongoing epidemics with H7N9 viruses, H7N9 viruses have been isolated from other species of wild land birds, including tree sparrow and wild pigeons (Zhao et al. 2014). These findings provide evidence that wild land birds may play an important role in the transmission of H7N9 subtype viruses. Here, we report the first isolation of H7N9 virus from magpie-robin in central China, in late 2015.
The location of the sampling site was in the suburb of Wuhan City adjacent to a small pool surrounding the villages. In these areas, birds are free ranging and mixing between chickens and domestic aquatic fowl (including ducks and geese) is common. The wild birds share the same area of water with domestic ducks. During October to December 2015, 230 fecal samples comprising 74 from tree sparrows (Passer montanus), 49 from magpie-robins (Copsychus saularis), 82 from pigeons (Columba), and 25 from Turtle Doves (Streptopelia turtur) were collected. The hosts for the samples were identified by mitochondrial analysis, as described previously (Cheung et al. 2009). For virus isolation, the fresh fecal samples were dissolved in PBS containing 0.1% bovine serum saline (BSA), 2 × 106 U/L penicillin G, 2 × 106 U/L amphotericin B, 250 mg/L kitasamycin, 0.5 × 106 U/L nystatin, 60 mg/L ofloxacin HCl and 400 mg/L streptomycin sulfate. The samples were centrifuged and the solution obtained was inoculated into 10-day-old SPF embryonated chicken eggs. After incubationat 37 ℃ for 72 h, the allantoic fluid was collected and its haemagglutination (HA) activity was tested. The HA positive allantoic fluids were confirmed by M gene specific reverse transcriptase-polymerase chain reaction (RT-PCR). The M gene positive samples were further analyzed by subtype specific RT-PCR. Only one fecal sample was positive for influenza virus and only H7 and N9 subtype-specific RNAs were detected.
To explore the genetic relationship between the strain we identified and other AIVs, the whole genome of the H7N9 virus, which named A/Magpie-Robin/Hubei/SZY300/2015 (SZY300/2015), was sequenced. Total RNA was isolated from individual samples using the RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. cDNA was synthesized using the Superscript Ⅲ First Strand Synthesis Kit (Invitrogen, Carlsbad, CA, USA). The Uni12 (5'-AGCAAAAGCAGG-3') primer was used for RT of the viral RNA genome. All segments were amplified using the Phusion TM HighFidelity PCR Kit (New England Biolabs, Ipswich, MA, USA). To sequence the full length genome, PCR was performed using universal primers for eight genes (Hoffmann et al. 2001). The nucleotide sequences from the virus were deposited into the GenBank database under accession nos. KY238127-KY238134.
A BLAST search showed that all 8 genes segments have high nucleotide identity (> 99%) with the poultry H7N9 viruses circulating in Jiangxi province around June 2014, indicating that the magpie-robin H7N9 virus isolated in late 2015 shares a common ancestor virus with the poultry H7N9 viruses circulating in other Chinese provinces in 2014 (Table 1). Phylogenetic inference was performed by RAxML 8.2.6 under the GTRGAMMA model with 1, 000 bootstrap replicates (Stamatakis 2014). All the sequences of H7N9 had been selected for phylogenetic analysis since 2013. The results confirmed that the 8 segments originated from the poultry H7N9 viruses circulating in Jiangxi province. In the phylogenetic tree of HA gene, the H7N9 virus of wild land bird origin clustered with the H7N9 viruses (A/chicken/Jiangxi/18515/2014-like) that circulated in poultry in mid-June 2014 in Jiangxi province (Supplementary Figure S1D). The H7N9 viruses were classified into the Yangtze River Delta lineage (YRD) and the Pearl River Delta lineage (PRD), with SZY300/2015 belonging to YRD (Wang et al. 2016) (Fig. 1). In the phylogenetic tree of neuraminidase (NA) gene, the H7N9 viruses show similar topology to the phylogenetic tree of HA gene, and the H7N9 virus from the wild land bird clustered with the H7N9 viruses circulating in Jiangxi and other Chinese provinces (Supplementary Figure S1F), indicating that they have common origin, and disseminated to diverse geographic locations. Phylogenetic tree analysis of the internal genes also indicated that this H7N9 virus is closely related to the H7N9 viruses (A/chicken/Jiangxi/18515/2014-like) that have circulated in Jiangxi province (Supplementary Figure S1). Thus, the H7N9 virus of wild land bird isolated from Hubei province in late 2015 perhaps originated from the poultry H7N9 viruses circulating in Jiangxi Province around June 2014.

Table 1. Sequence homologies of the whole genome of the wild land bird origin H7N9 virus, with nucleotide sequences available in GenBank.
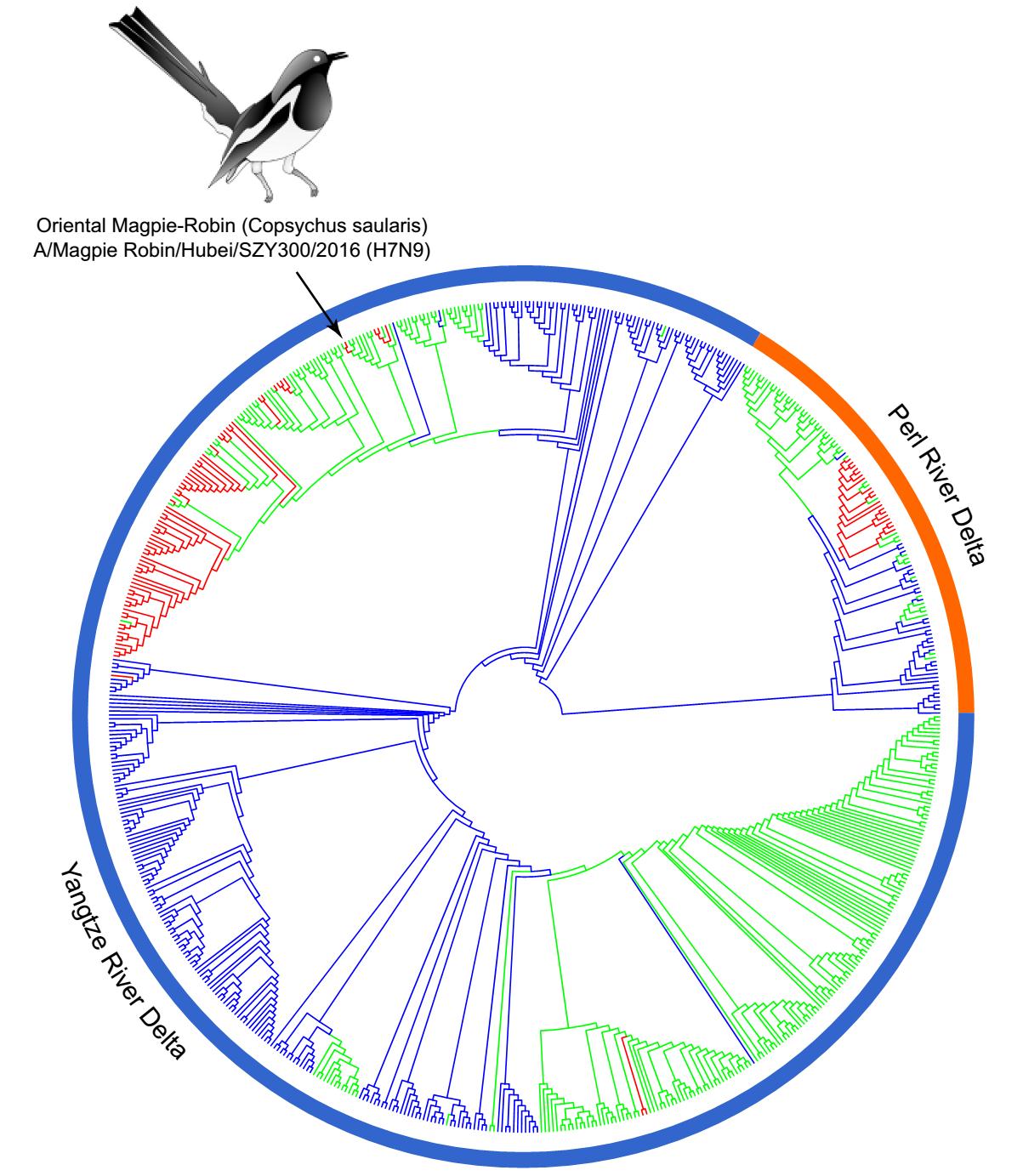
Figure 1. Maximum-likelihood phylogenetic trees of the HA (positions 1-1683) gene of H7N9 influenza virus. Phylogenetic inference was performed by RAxML 8.2.6 under the GTRGAMMA model with 1000 bootstrap replicates. The virus obtained from the present study is marked by arrow.
Amino acid sequence analysis identified G186V and Q226L substitutions in the HA protein (H3-numbering), indicating that the virus had a potential to bind to humanlike receptors (Gambaryan et al. 2012; Xiong et al. 2013). The PEIPKGR/G amino acid motif at the HA cleavage site of the H7N9 isolate contains two basic amino acids, which is a typical characteristic sequence for low pathogenic AIVs (Supplementary Table S1). An amino acid deletion of 69-73 residues in length was also found in the NA gene (N2). Amino acid 294R was maintained in neuraminidase genes, indicating its susceptibility to oseltamivir (Supplementary Table S1) (Wu et al. 2013). Analysis of the internal proteins of this isolate indicated that, in the polymerase complex, no amino acid substitutions were found in the key sites related to host tropism and virus replication capacity (e.g., E158G, E627K and D701N in the PB2 protein, Y436H in the PB1 protein, and T515A in the PA protein), suggesting its low pathogenicity towards mammals (Supplementary Table S1) (Bi et al. 2015). However, an S31N mutation in the matrix 2 protein confers adamantanes resistance to the H7N9 virus from the wild land bird (Supplementary Table S1) (Pinto et al. 1992).
The H7N9 AIV isolated from a magpie-robin in Hubei province has high sequence similarity with the H7N9 viruses isolated from poultry in Jiangxi province. This indicated that virus transmission might have occurred between these two regions. Two transmission routes are possible: (1) Wild land birds in Jiangxi province acquired the H7N9 virus from Jiangxi poultry and then spread it to the wild land bird in Hubei province; (2) H7N9 virus infected poultry from Jiangxi province were transported to Hubei province and the virus then spread to poultry in Hubei, which subsequently transmitted it to wild land birds in Hubei province. However, other factors related to the transmission of H7N9 from Jiangxi to Hubei cannot be excluded.
In previous reports, H5N1 influenza viruses were isolated from magpie-robin in Hong Kong and a serologic survey also showed the presence of serum neutralization antibody against H5N1 in these birds (Govorkova et al. 2013; Siengsanan-Lamont et al. 2011). However, there are no reports documenting the isolation of other influenza virus subtypes from magpie-robins. Our isolation of the H7N9 virus from a magpie-robin merits attention. However, we cannot ascertain whether the virus is stable in magpie robins because of the small sample size of this study. After all, wild bird surveillance of novel H7N9 viruses requires large sample sizes. Magpie-robins are abundant and widely distributed in Hubei and they frequently come into contact with humans and poultry. The presence of Magpie-robin-derived-H7N9 has a potential to increase opportunities for spread to domestic farms and even to humans. Additionally, the sample sites used in the present study were around densely populated areas, thereby highlighting the seriousness of the threat posed by this novel subtype.
In summary, we isolated an H7N9 virus from a wild Magpie-robin in late 2015 from Hubei and genetic analysis showed that it shares a common ancestor with the poultry H7N9 viruses from Jiangxi in 2014. This makes the control of this H7N9 subtype virus challenging. Thus, it is important to continue epidemiological monitoring to determine whether the H7N9 virus has become endemic in wild land birds.
-
This research is supported by grants from the National Natural Science Foundation of China (31400787, 31570026), Hubei Provincial Natural Science Foundation of China (2016CFC743), Wuhan Youth Science and technology plan (2016070204010104) and Special Project of Ministry of Science and Technology (2013FY113500).
-
The authors declare that they have no conflicts of interest.
-
The manuscript did not use animal and human subjects since fecal samples were collected in the environment.
Conflict of Interest
Animal and Human Rights Statement
-

Figure S1. Figure S1. Maximum-likelihood phylogenetic trees of the coding sequences of the eight segments. A) PB2, B) PB1, C) PA, D) HA, E) NP, F) NA, G) M, H) NS. Node labels indicate bootstrap values. Colored tip and nodes are: blue, H7N9 viruses isolated in 2013; green, H7N9 viruses isolated in 2014; red, H7N9 viruses isolated in 2015.

Table S1. Table S1 Molecular characterization of H7N9 at representative sites*

 DownLoad:
DownLoad: