












HTML
-
Because of its high contagiousness, nearly everyone has come into contact with varicella-zoster virus (VZV) before adulthood, either with wild strains, which may cause chickenpox/varicella upon primary exposure, or with attenuated strains (e.g. Oka strains), which are used as varicella vaccines. Unfortunately, all of these viruses can lurk in the sensory ganglia and reactivate as shingles/zoster by taking advantage of weakened cellular immune responses caused by aging (Garnett and Grenfell 1992; Krause and Klinman 2000; Arruti et al. 2017; Breuer 2018). As antiviral drugs and pain relievers can alleviate only some of the associated symptoms, vaccine strategies have become the first choice to address VZV infection.
Since its approval by the FDA in 1995, the Oka strain developed by Michiaki Takahashi has been used worldwide as a varicella vaccine at a dose of approximately 1000–10, 000 plaque-forming units (PFU) (Zhu et al. 2016). Herpes zoster after vaccination implied that there is a risk of latent infection similar to wild strains and a subsequent risk of herpes zoster (Krause and Klinman 2000; Uebe et al. 2002; Moodley et al. 2019). Interestingly, booster immunization with the Oka strain at a dose as high as 20, 000 PFU (i.e., Zostavax developed by Merk in 2005) could be used as a herpes zoster vaccine, mainly due to its ability to boost pre-existing cellular immune responses upon primary exposure or vaccination. In addition to difficulties in manufacturing and maintaining the necessary high titers of this live-attenuated virus, the protective rate declines from 70% for people aged 50–59 years to less than 38% for people older than 70 years, which may be attributed to senescence of the immune system and reduced efficiency in boosting VZV-specific cell-mediated immunity (CMI) (Oxman et al. 2005; Gilderman et al. 2008; Asada 2019).
Shingrix is a subunit zoster vaccine developed by GlaxoSmithKline (GSK), with the conserved VZV glycoprotein E (gE) as the antigen and the AS01B adjuvant system (Dendouga et al. 2012). As one of the most abundant glycoproteins, gE plays important roles in virus replication and transmission between ganglia cells (Berarducci et al. 2006; Malavige et al. 2008). With the inclusion of AS01B to enhance CMI targeting the extracellular domain of gE expressed in Chinese hamster ovary (CHO) cells, Shingrix showed protective rates of 96.6%, 97.4% and 91.3% for people aged 50–59, 60–69 and older than 70 years, respectively (Haumont et al. 1996; Leroux-Roels et al. 2012; Chlibek et al. 2013; Cohen 2015; Cunningham et al. 2016; Cunningham et al. 2018). In addition, as a subunit vaccine, it also showed good safety and efficacy in people with immune compromising diseases, including HIV carriers, autologous hematopoietic cell transplant recipients, renal transplant recipients, patients with solid tumors, etc. (Stadtmauer et al. 2014; Berkowitz et al. 2015; Levin et al. 2017; Bastidas et al. 2019; Vink et al. 2019; Cheng et al. 2020). One of the key components of AS01B (i.e., QS21) for inducing CMI is a fraction of an extract from the bark of Quillaja saponaria. Because it is a polysaccharide mixture, synthesis of QS21 is infeasible (Upadhyay et al. 2018). The limited distribution of Q. saponaria around the globe and the strict quality control during the extraction processes of QS21 have made Shingrix very expensive (approximately 150–200 USD per dose). In addition, mixing strategies that are helpful for the stability of this vaccine also cause a higher cost of production.
In our previous reports, we demonstrated the synergistic effects of polyinosinic-polycytidylic acid (Poly I:C) and CpG oligodeoxynucleotide (CpG ODN) as adjuvants to enhance CMI induced by the human papilloma virus (HPV) E7 epitope vaccine in a TC-1-grafted tumor model (Liu et al. 2018a). In addition, we have proven that encapsulation of the natural phosphodiester CpG ODN in poly(lacticco-glycolic acid) (PLGA) nanoparticles showed efficacy comparable to that of phosphorothioate CpG ODN with the same sequences at the same doses but was much more economical (Liu et al. 2018b). Considering that the key success criterion of the subunit zoster vaccine is the ability to enhance CMI to specific antigens, which is similar to the principle followed for tumor vaccines (Levin et al. 2018b; Liu et al. 2018a), we designed a series of nanoparticles based on PLGA with nucleic acids as adjuvants and tested their efficacy in enhancing CMI targeting VZV-gE, which may offer a much more economical strategy for the development of zoster vaccines.
-
The composition of the designed vaccines is shown in Table 1.
Vaccine group gE (μg) CpG (μg) Poly I:C (μg) In Out BW006 2395 ① gE 10 ② gE + CpG + Poly I:C 10 5 5 25 ③ gE + Alum 10 ④ gE + Freund's 10 ⑤ DOTAP-IN 10 5 5 25 ⑥ DOTAP-HALF 5 5 5 5 25 ⑦ DOTAP-OUT 10 5 5 25 ⑧ PLGA 10 5 5 25 ⑨ DOPC 10 5 5 25 ①-④ are mixtures, and ⑤-⑨ are nanoparticles all contain gE+CpG+Poly I:C. For DOTAP-IN (⑤), gE only exists in the inner water phase of the (water/oil/water) emulsions; for DOTAP-OUT (⑦), gE only exists in the outer water phase; for DOTAP-HALF (⑥), gE exists in both water phases. Table 1. Composition of each dose of the designed vaccines.
The extracellular domain of gE expressed in CHO cells was supplied by AtaGenix Laboratory (Wuhan, China). Phosphodiesters CpG ODN BW006 (class B, 5'-tcg acg ttc gtc gtt cgt cgt tc-3') and 2395 (Class C, 5'-tcg tcg ttt tcg gcg c:gc gcc g-30) were synthesized and purified via high-performance liquid chromatography (HPLC) by Sangon Biotech Co., Ltd. (Shanghai, China). Low molecular weight (LMW) poly I:C was purchased from InvivoGen (tlrl-picw-250), alum adjuvant was purchased from Thermo Fisher (77161), and Freund's adjuvant was purchased from Merck (F5581 & F5506). For groups ① gE, ② gE + CpG + Poly I:C, ③ gE + Alum and ④ gE + Freund's, vaccines were formulated by mixing these agents in phosphate-buffered saline (PBS, pH 7.4) with a pipettor.
Nanoparticle vaccines based on PLGA were formulated with a double-emulsion (W/O/W) solvent evaporation method (Liu et al. 2016a). Briefly, 1 mL of dichloromethane (DCM, Merk, 270997) containing 30 mg of PLGA (Merk, 719897) without (for group ⑧ PLGA) or with 6.5 mg of the cationic lipid 1, 2-dioleoyl-3-trimethylammonium-propane (DOTAP, Merk, 890890P, for groups ⑤-⑦, i.e., DOTAPIN/HALF/OUT) or 6.5 mg of the neutral lipid 1, 2-dioleoylsn-glycero-3-phosphocholine (DOPC, Merk, 850375P for group ⑨ DOPC) were mixed with PBS solution containing gE, CpG ODNs and Poly I:C and sonicated over an ice bath with a microtip probe sonicator (Sonics & Material Inc., Connecticut, USA) at a power of 30 kW for 1 min. After the addition of a secondary aqueous phase, i.e., 5 mL of 2% w/v polyvinyl alcohol (Merk, P8136) in deionized water, the primary water-in-oil (W/O) emulsion was further sonicated for 5 min to obtain a secondary emulsion (W/O/W) and agitated overnight with a magnetic stir bar at room temperature to evaporate the DCM completely. The final product was collected by centrifugation at 21, 000 rpm for 30 min, washed with distilled water and lyophilized. For groups containing gE on the surface of nanoparticles (groups ⑥ DOTAP-HALF and ⑦ DOTAP-OUT), aqueous solutions containing gE were gently stirred with DOTAP-PLGA nanoparticles overnight at 4 ℃ for adsorption. The gE-ad-sorbed nanoparticles were washed three times with distilled water via centrifugation at 21, 000 rpm for 30 min and lyophilized.
With PLGA as a control, the prepared vaccines were dissolved in 0.1 mol/L NaOH and 0.1% sodium dodecyl sulfate (SDS) overnight at room temperature. Loaded gE was detected with a bicinchoninic acid (BCA) protein assay kit (Beyotime, P0012). Loaded nucleic acid adjuvants were detected with a Quant-iT OliGreen ssDNA Reagent Kit (Thermo Fisher, O11492). Before immunization, the nanoparticles were suspended in PBS and observed under a transmission electron microscope (TEM, Hitachi Ltd., Tokyo, Japan) (Liu et al. 2015).
-
Female specific pathogen-free (SPF) C57BL/6N mice at 5 weeks of age (16–20 g) were supplied by Vital River Laboratory Animal Technology Ltd. (Beijing, China) and maintained under SPF conditions at the Central Animal Services of Medical Biology of the Chinese Academy of Medical Sciences, Peking Union Medical College. The animals were randomly divided into 10 groups with 8 mice in each group (N = 8). The mice were immunized intramuscularly in the thigh muscle three times with 50 μL of immunogen at 2-week intervals. Blood samples (via heart puncture) and spleens were collected 2 weeks after the final immunization. After clotting at 4 ℃ overnight, serum was collected after centrifugation at 3000 rpm for 10 min.
-
gE (2 μg/mL) was coated on 96-well plates (Corning, USA) at 4 ℃ overnight. After blocking with 5% (w/v) skim milk at 37 ℃ for 1 h, the plates were incubated with serial dilutions of mouse sera at 37 ℃ for 1 h. Bound antibodies were detected with goat-anti-mouse IgG-HRP (horseradish peroxidase) conjugate (1:5000, Bio-Rad, 5178-2504) as a secondary antibody. Ten minutes after the addition of the substrate 3, 3', 5, 5'-tetramethylbenzidine (TMB, BD, 555214), 1 mol/L phosphoric acid was added to terminate the reaction. Absorbance at 450 nm was detected with a spectrophotometer (BioTek, USA).
-
Spleens were dispersed with a 70 μm cell strainer (BD, USA), and splenocytes were separated with a Ficoll-Isopaque density gradient centrifugation kit (Multi Sciences, 70-LSM01). After calculation, splenocytes were suspended in Roswell Park Memorial Institute (RPMI) 1640 medium (Thermo Fisher, 61870127) with 10% v/v fetal bovine serum (FBS, Biological Industries, 04-001-1ACS) and penicillin-streptomycin (Thermo Fisher, 15140122) at a final concentration of 1 × 107 cell/mL. Then, 100 μL of cells was added into each well of a 96-well plate (Corning, USA) for further analysis.
-
Splenocytes from 8 mice in each group were further divided randomly into two subgroups, one for the detection of antigen-specific IL-2-producing cells and the other for the detection of IFN-γ-producing cells. With ELISPOT assay kits (551076 for IL-2 and 551083 for IFN-γ, both from BD) and according to the manufacturers' protocol, tests were carried out as follows: 1 × 106 cells were incubated with 10 μg/mL gE, 10 μg/mL pooled peptides or a mixture of 5 μg/mL gE and 5 μg/mL pooled peptides for 16-18 h at 37 ℃ with 5% CO2. Pooled peptides were those T cell epitopes of gE predicted with tools from the Immune Epitope Database at http://tools.iedb.org and matched with previous reports (Garcia-Valcarcel et al. 1997). Four of these peptides (corresponding to sequences: gE (11-30): VLMGFGIITGTLRITNPVRA; gE (71-90): SRKAYDHNSPYIWPRNDYDG; gE (91-110): FLENAHEHHGVYNQGRGIDS; gE (106-125): GRIDSGERLMQPTQMSAQED) were selected and synthesized by GL Biochem Co., Ltd (Shanghai, China) with purity ≥ 95% to stimulate gE-specific T cell responses.Spots were counted with an ELISPOT reader system (Autoimmun Diagnostika GmbH, Germany) after immunoimaging.
-
A total of 1 × 106 splenocytes were incubated with 5 μg/mL gE or 5 μg/mL pooled peptides together with 5 μL/well FastImmune (BD, 347690) at 37 ℃ with 5% CO2 for 2 h, and brefeldin A (Biolegend, 420601) was added and incubated overnight under the same conditions to block cytokine release. After washing with staining buffer (Biolegend, 420201), 5 μg/mL CD16/32 antibodies (Biolegend, 101326) were added and incubated at 4 ℃ for 10 min to block nonspecific binding of Fc. After incubation with FITC antimouse CD4 antibodies (Biolegend, 100405) and PerCP/ Cyanine5.5 anti-mouse CD8a antibodies (Biolegend, 100733) at 4 ℃ for 30 min, the cells were fixed with fixation buffer (Biolegend, 420801) in the dark at room temperature for 20 min. After washing with permeabilization wash buffer (Biolegend, 421002), APC anti-mouse IL-2 antibodies (Biolegend, 503809) and PE anti-mouse IFN-γ antibodies (Biolegend, 505807) were added and kept in the dark at room temperature for 30 min. After one wash with permeabiliza- tion wash buffer and one wash with PBS, the cells were analyzed with a BD Accuri C6 Flow Cytometer and FlowJo software (BD, USA).
-
Significant differences among experimental groups were analyzed using one-way analysis variance (ANOVA) followed by Dunnett's multiple comparisons test, compare the mean of each group with the mean of PLGA group as control (GraphPad Prism 7.0 software, GraphPad Software Inc., La Jolla, CA, USA). *P < 0.05. **P < 0.01. ***P < 0.001. ****P < 0.0001. ns, no significant difference.
Vaccine Preparation and Characterization
Immunization of Mice
Enzyme-Linked Immunosorbent Assay (ELISA) for gE-Specific IgG
Preparation of Splenocytes
Enzyme-Linked Immunospot Assay (ELISPOT)
Flow Cytometry
Statistical Analysis
-
The compositions of the prepared nanoparticle vaccines are shown in Table 2, and the data were obtained as the average of three measurements. To test gE presented on the surface of group ⑥ DOTAP-HALF, a portion of the semifinished nanoparticles (after gE encapsulation but before gE absorption) were collected and deducted from the finished nanoparticles. The final contents of CpG ODN BW006, 2395 and Poly I:C were calculated by their designed proportion in Table 1 and the total nucleic acid contents tested.
Vaccine group gE (mg) CpG (mg) Poly I:C (mg) In Out BW006 2395 ⑤ DOTAP-IN 0.53 0.19 0.19 0.98 ⑥ DOTAP-HALF 0.32 0.30 0.30 0.30 1.53 ⑦ DOTAP-OUT 0.61 0.27 0.27 1.38 ⑧ PLGA 0.55 0.21 0.21 1.07 ⑨ DOPC 0.5 0.17 0.17 0.86 Table 2. Composition of 50 doses of the nanoparticle vaccines.
Two typical forms of nanoparticles are shown under TEM. One has a diameter of approximately 200 nm (shown by the arrows hit to the right in Fig. 1), which is the approximate diameter of VZV, i.e., 180–200 nm, and is mainly present among nanoparticles without cationic lipids (groups PLGA and DOPC). Nanoparticles with cationic lipids (groups DOTAP-IN, DOTAP-HALF and DOTAP-OUT) are mainly composed of particles with much smaller diameters (shown by the arrows hit to the left in Fig. 1).
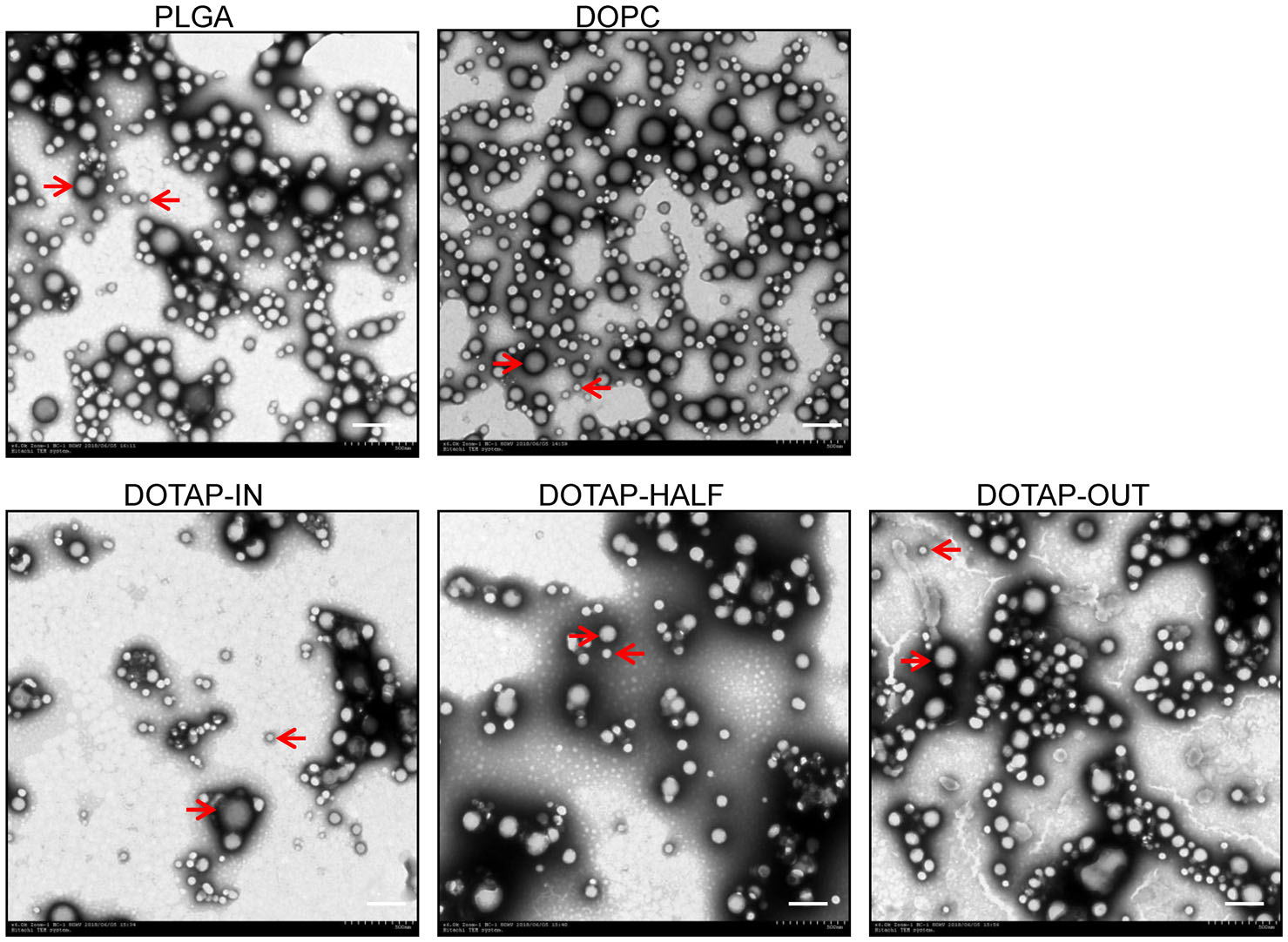
Figure 1. Nanoparticles under a transmission electron microscope. Particles with diameters of approximately 200 nm are shown by the arrows pointing to the right, which are mainly present among nanoparticles without cationic lipids (groups PLGA and DOPC). And particles with diameters of approximately 50 nm are shown by the arrows pointing to the left, which are mainly present among nanoparticles with cationic lipids (DOTAP). Scale bar, 250 nm.
-
Two weeks after the last immunization, blood of tested C57BL/6N mice was collected (Fig. 2A). Serum samples from the mice in group PLGA showed the highest gEspecific IgG titers (592981) among the mice immunized with nanoparticles; these titers were only lower (P = 0.4339) than those of the mice in the Freund's adjuvanted group (816702) with pipettor-mixed vaccines. The titer was much higher than that of the DOTAP-HALF nanoparticle group (140972, P = 0.0077) or the group treated with the pipettor-mixed vaccine with the same antigens and nucleic acid immune stimulators (gE + CpG + Poly I:C group in Fig. 2B, 214633, P = 0.0370).
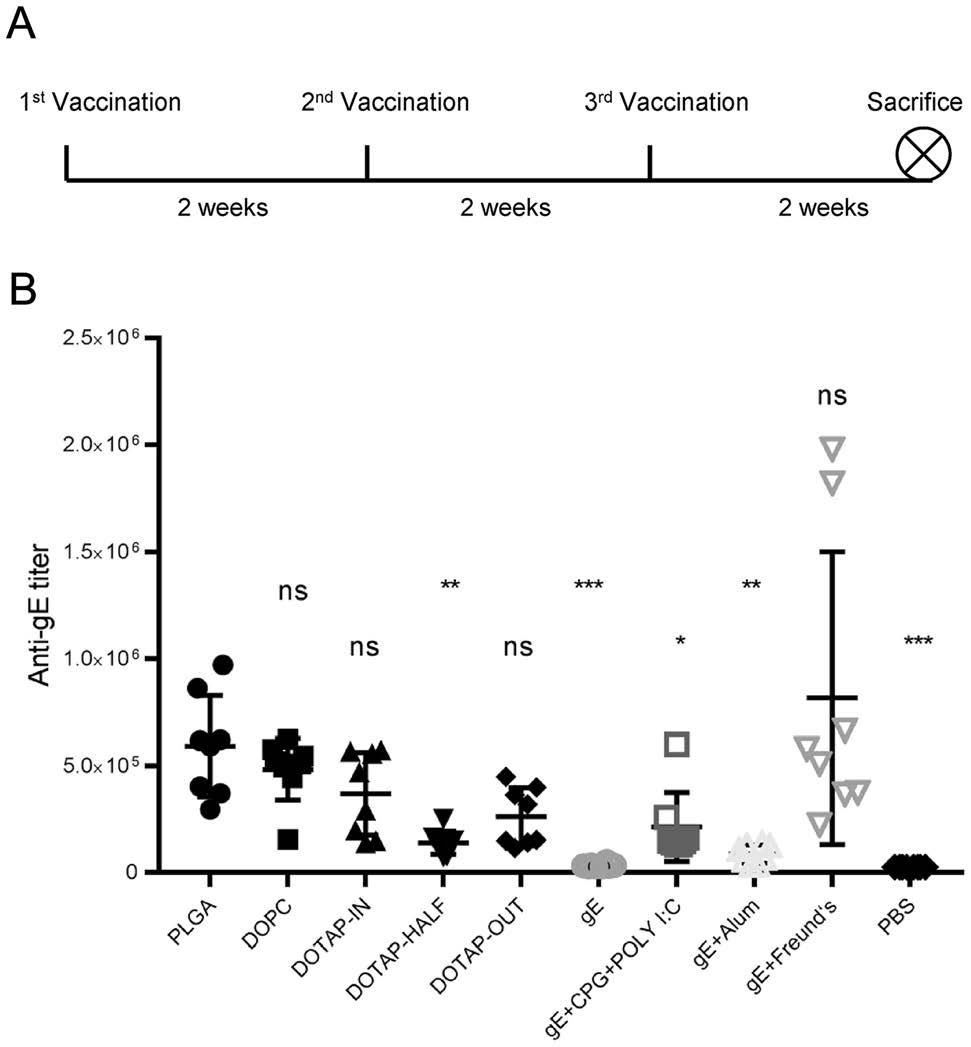
Figure 2. Immunization schedules (A) and gE-specific IgG titers (B). Anti-gE titers of each group were compared with that of the PLGA group. *P < 0.05. **P < 0.01. ***P < 0.001. ns, no significant difference.
-
Similar numbers of IFN-γ-producing splenocytes were detected in each group of mice immunized after stimulation with 10 μg/mL gE (Fig. 3A) or with 10 μg/mL pooled peptides (Fig. 3B). This similarity was also observed for IL-2-producing splenocytes (Fig. 3D, 3E). A mixture of 5 μg/mL gE and 5 μg/mL pooled peptides as stimulants showed no prominent effects on the numbers of either IFN-γ-producing (Fig. 3C) or IL-2-producing (Fig. 3F) splenocytes.
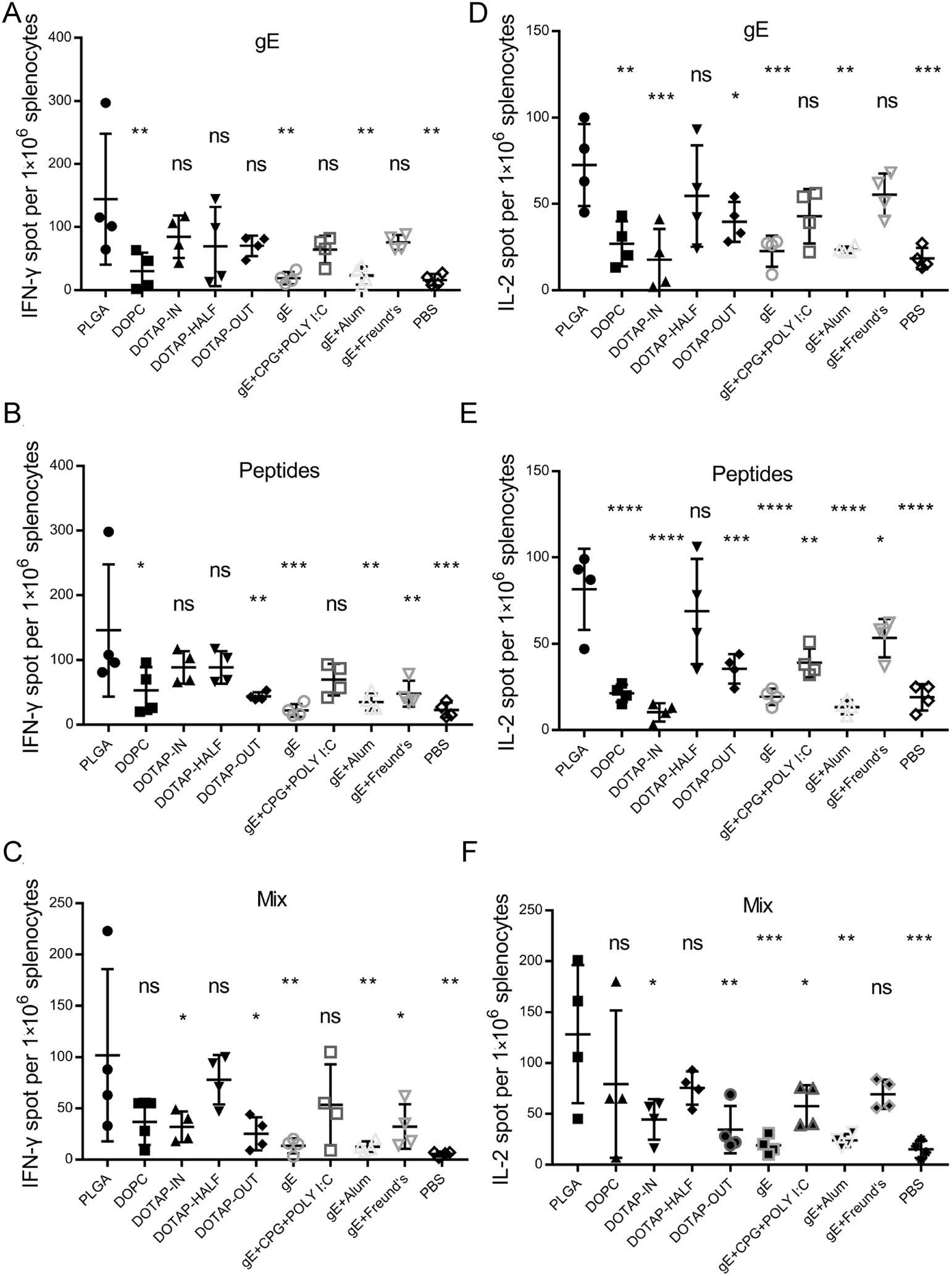
Figure 3. Enzyme-linked immunospot assay (ELISPOT) of splenocytes. A–C IFN-γ-producing splenocytes after stimulation with gE, pooled peptides, or mixtures of gE and pooled peptides. D–F IL-2-producing splenocytes after stimulation with gE, pooled peptides, or mixtures of gE and pooled peptides. Spot numbers of each group were compared with that of the PLGA group.*P < 0.05. **P < 0.01. ***P < 0.001. ****P < 0.0001. ns, no significant difference.
The PLGA group showed both the most IFN-γ-producing splenocytes and the most IL-2-producing splenocytes among all of the immunized groups (including nanoparticle vaccines and pipettor-mixed vaccines). While loading of antigens and nucleic acid adjuvants with PLGA promoted a limited increase in IFN-γ-producing splenocytes (PLGA group versus gE + CpG + Poly I:C group in Fig. 3A–3C), nanoparticle formation was quite helpful for increasing in IL-2-producing splenocytes (PLGA group versus gE + CpG + Poly I:C group in Fig. 3D–3F). While encapsulation of PLGA-nucleic acid adjuvant systems in either neutral (group DOPC) or cationic lipids (groups with DOTAP) reduced not only IFN-γ-producing but also IL-2-producing splenocytes to different extents, presentation of gE in nanoparticles that contained cationic lipids could reduce this disadvantage to a certain extent. In contrast to the DOTAP-IN and DOTAP-OUT groups, the DOTAP-HALF group showed higher IFN-γ- and IL-2-producing splenocytes but still fewer than the PLGA group after stimulation with different combinations, but no significant differences were detected (Fig. 3).
-
After stimulation with 5 μg/mL gE (Fig. 4A, 4C) or 5 μg/mL pooled peptides (Fig. 4B, 4D), higher proportions of antigenspecific CD4+ (Fig. 4A–4B) and CD8+ (Fig. 4C–4D) T cells (including those producing IFN-γ alone, those producing L-2 alone and those producing both IFN-γ and IL-2) were detected in splenocytes of both the mice immunized with PLGA nanoparticles and the mice immunized with DOTAPHALF nanoparticles. Although there was no significant difference in the proportion of antigen-specific CD8+ T cells among splenocytes between the PLGA and DOTAP-HALF nanoparticle-immunized mice, much higher proportions of gE-specific (P = 0.0106, Fig. 4A) and peptide-specific (P = 0.0305, Fig. 4B) CD4+ T cells were detected in the splenocytes of the PLGA nanoparticle-immunized mice.
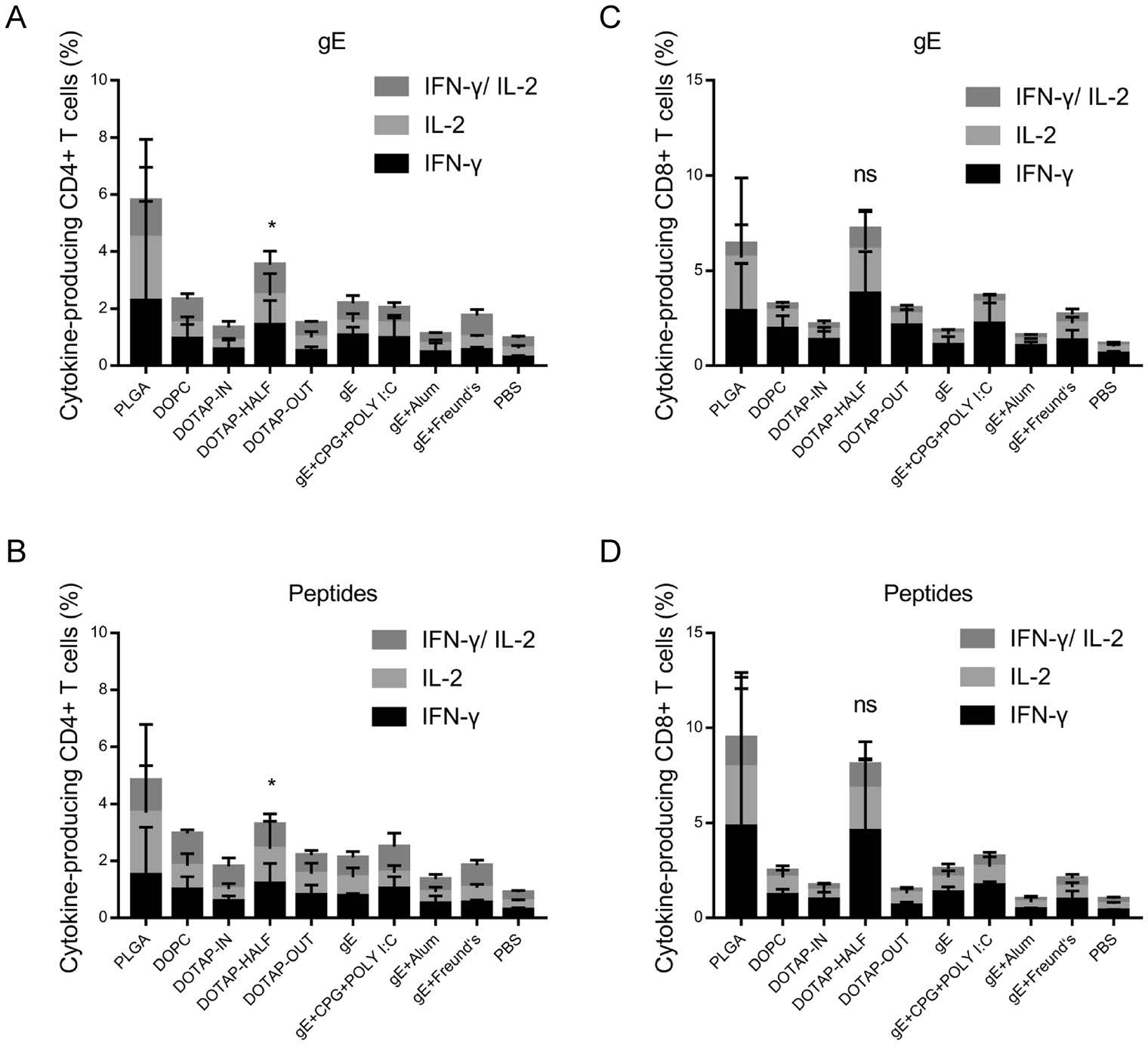
Figure 4. Flow cytometry assay for antigen-specific T cells. A–B Proportion of IFN-γ- and IL-2-producing CD4+ T cells among splenocytes after stimulation with gE or pooled peptides stimulation. C–D Proportion of IFN-γ- and IL-2-producing CD8+ T cells among splenocytes after stimulation with gE or pooled peptides. Percentage of cytokine-producing T cells of each group was compared with that of the PLGA group. *P < 0.05. ns, no significant difference.
Nanoparticle Vaccine Characterization
gE-Specific IgG Titers
More gE-Specific IFN-γ- and IL-2-Producing Splenocytes were Detected in the PLGA Nanoparticle-Immunized Group than in the Other Groups
Higher Proportions of gE-Specific CD4+ and CD8+ T Cells were Detected in the PLGA Nanoparticle-Immunized Group than in the Other Groups
-
Although in vitro experiments showed that reactivation of the Oka strain is impaired, clinical reports showed that reactivation of Oka strain is well established and implied a subsequent risk of herpes zoster, which pose greater threat to immune compromised patients (Uebe et al. 2002; Sadaoka et al. 2016; Levin et al. 2017; Levin et al. 2018b; Moodley et al. 2019). In addition, the strict conditions required during transportation of the zoster vaccine based on the Oka strain and the rapid decline of protective efficacy for people older than 70 years need to be solved (Oxman et al. 2005; Gilderman et al. 2008).
Subunit vaccines are good choices to avoid the latency of viruses from live-attenuated vaccines, but their immunity should be strengthened further. As the component essential for the replication and transmission of VZV, the conserved gE protein, which contains potential neutralization epitopes and T cell epitopes, is an ideal antigen for VZV subunit vaccines (Garcia-Valcarcel et al. 1997; Berarducci et al. 2006; Malavige et al. 2008; Zhu et al. 2016). After immunization of mice with one linear B cell epitope of gE presented on hepatitis B core virus-like particles (VLPs), the collected serum could neutralize VZV infection in vitro (Zhu et al. 2016; Wu et al. 2017). Unlike VLPs, which suffer from limited engineer ability for enhancing antigen immunity, synthetic particulate systems are highly tunable (Rosenthal et al. 2014; Jain et al. 2015). Codelivered with nucleic acid adjuvants as immune potentiators, particles based on PLGA and prepared with nanotechnology (Fig. 1) showed much higher gE-specific IgG titers (P = 0.0012) than pipettor-mixed vaccines with alum as an adjuvant (Fig. 2), which showed promise for use as subunit varicella vaccines. With liposomes instead of nanoparticles as delivery systems and the combination of QS21 and monophosphoryl lipid A (MPL) as immune potentiators, AS01B adjuvant systems induced gE-specific IgG titers approximately 2 times higher than those induced by alum adjuvant systems (Dendouga et al. 2012). Although a subunit vaccine based on AS01B-adjuvanted gE, i.e., Shingrix, has been on the market to prevent shingles, there are no clinical reports concerning whether this vaccine can be used as a safer varicella vaccine. One rational explanation may be that it is not cost-effective to replace the current live-attenuated varicella vaccine on the market with Shingrix, which is in short supply and sold at much higher prices.
In addition to the limited supply of QS21, another factor increasing the cost is that Shingrix is supplied as separate vials of lyophilized gE and liquid liposomal adjuvants to avoid damaging the physiochemical characteristics of AS01B during the drying and rehydration process. Cationic lipids are helpful in increasing the efficacy of similar liposomal adjuvant systems during the drying and rehydration process, but there are reports concerning side effects related to the cytotoxicity of cationic lipids (Romoren et al. 2004; Liu et al. 2016a; Cui et al. 2018; Wui et al. 2019). To our surprise, nanoparticles loaded with nucleic acid adjuvants without cationic lipids (the PLGA group in Figs. 3, 4) are more potent in inducing CMI than the same particles coated with cationic lipids (the DOTAP group in Figs. 3, 4). This reminds us that although both cationic lipids and nucleic acid adjuvants are critical to induce antigen cross-presentation, which is helpful to induce CMI targeting vaccine antigens, interference between these two components may neutralize their efficacy (Maurer et al. 2002; Bode et al. 2011; Gwak et al. 2016; Gao et al. 2017). This neutralization may be partly caused by the bonding effect of coated lipids, which blocks the release of encapsulated antigens and nucleic acid immune stimulators (PLGA versus DOPC in Figs. 3, 4). As shown in the DOTAP-HALF group (in Figs. 3, 4), an adequate amount of antigen initially exposed on the outside of the nanoparticles and long-term release of encapsulated antigens are most efficient in enhancing CMI among different forms of nanoparticles containing cationic lipids, which may also occur in PLGA particles without bonding of coating lipids (Liu et al. 2016b).
While CD4+ T cells played pivotal roles in the control of simian varicella virus infection in a nonhuman primate model, both VZV-specific CD4+ and CD8+ T cells play important roles in recovery from varicella and control of active herpes zoster (Weinberg and Levin 2010; Haberthur et al. 2011; Steain et al. 2014). Consistent with the higher protection rate of Shingrix, recipients of Shingrix showed approximately 10 times more gE-specific CD4+ and CD8+ T cells than recipients of Zostavax (Levin et al. 2018b). Notably, gE-specific Th1 CD4+ T cells are adopted more frequently than CD8+ T cells as good indicators for the potential of zoster vaccines in animal experiments and clinical trials (Dendouga et al. 2012; Laing et al. 2015; Cunningham et al. 2018). This may be attributed to two reasons. The first is that shortly after immunization, MHC Ⅱ dendritic cells are increased to a high level and are more efficient in presenting antigens to CD4+ T cells than CD8 T cells (Didierlaurent et al. 2014). The second is that unlike antigen-specific CD4+ T cells, after elimination of infected pathogens or immunized antigens, memory CD8+ T cell decrease very quickly (Varga et al. 2001; Seder and Ahmed 2003; Weinberg et al. 2018). In contrast to previous reports of liposome-based subunit zoster vaccines in animal experiments, PLGA-based nanoparticles together with nucleic acid immune stimulators are powerful tools for producing not only gE-specific CD4+ T cells but also gE-specific CD8+ T cells (Fig. 4C, 4D) (Dendouga et al. 2012; Wui et al. 2019). The higher proportion of gE-specific CD8+ T cells in our experiments may be attributed to the antigen cross-presentation ability of nucleic acid adjuvants and tight schedules for the sacrifice of immunized mice, which may avoid deceases in CD8+ T cells. According to our previous experience with tumor models, if immunized mice were sacrificed one month later (which is closer to the sacrifice time interval reported for liposome-based subunit zoster vaccines) rather than two weeks later, antigenspecific CD8+ T cells decreased to a low level (Liu et al. 2018a; Liu et al. 2018b). Though whether these CD8+ T cells are helpful for the protective efficacy of zoster vaccines is still to be validated, they show little potential safety risk because cytolytic CD8+ T cells that infiltrate around ganglia with reactivating VZV do not induce neuronal apoptosis (Steain et al. 2014).
Due to the lack of high-quality Oka strain-based vaccines with high titers, such as Varilrix from GSK and Zostavax from Merk, we did not adopt the VZV-primed mouse model to evaluate the potency of our PLGA nanoparticles as zoster vaccines (Dendouga et al. 2012; Wui et al. 2019). Instead, we administered three doses of the vaccine (Fig. 2A), comparable to the immunization schedule of one dose of live VZV before two doses of subunit vaccine that was reported for the development of Shingrix in animal experiments (Dendouga et al. 2012). Although priming with VZV will not result in latency and reactivation in mice, there are reports that viral DNA could be found 1 month after inoculation, which implied probable replication of VZV in mice, maybe at a low level (Wroblewska et al. 1993; Mahalingam et al. 2010). The persistent existence of VZV in vivo means persistent stimulation of the immune system and a comparably higher basal level of VZV-specific immunity (Levin et al. 2018a). Although the boosting effects of the first dose of nanoparticle vaccines may not be as potent as those of lasting VZV viruses in vivo, effective induction of CMI by these nanoparticle vaccines implied comparable or perhaps better potency of those vaccines than observed in VZV-primed mouse models.
In conclusion, encapsulation of VZV-gE protein as an antigen with nucleic acids, including Poly I:C and phosphodiester CpG ODNs, as immune stimulators in nanoparticles based on PLGA induced not only high IgG titers which showed promise for use as a subunit varicella vaccine that will not cause virus latency or a potential risk of zoster, but also high CMI which showed promise for use as a zoster vaccine. These particles are stable during the lyophilization and rehydration process, and all of the components included in the formulations are safe and economical. These characteristics make these nanoparticles a potentially safe and economical varicella and zoster vaccine candidate.
-
This work was financially supported by the CAMS Initiative for Innovative Medicine (Grant Number 2017-I2M-3-022), Central basic scientific research in colleges and universities (Grant Number 3332019162), the National Natural Science Foundation of China (Grant Number 81503117) and the Foundation for Studying Abroad from the China Scholarship Council (Grant Number 201808110121).
-
YW and JQ performed the experiments and analyzed the data. HC helped to perform part of the experiments. CL designed the study, drafted and finalized the manuscript.
-
The authors declare that they have no conflict of interest.
-
The animal experiments were approved by the Ethics Committee of Animal Care and Welfare of the Institute of Medical Biology, Chinese Academy of Medical Sciences and Peking Union Medical College (Permit Number: SCXK (dian) K2017-0002), in accordance with the animal ethics guideline of the Chinese National Health and Medical Research Council and the Office of Laboratory Animal Management of Yunnan Province, China. All efforts were made to minimize animal suffering.

 DownLoad:
DownLoad: