














HTML
-
Hepatitis E virus (HEV) is one of the commonest global causes of hepatitis (Aggarwal and Naik 2009). Seven closely related HEV genotypes (HEV-1–7) have been identified, but only four major HEV genotypes cause human infection, each of which has different epidemiological features (Woo et al. 2016). HEV genotype 1 had been isolated in Asia and Africa and genotype 2 was originally identified in Mexican and some African isolates (Kamar et al. 2012). Genotype 3 was initially isolated from human patients in the United States and has since been detected on most continents, including Europe (Vina-Rodriguez et al. 2015; Adlhoch et al. 2016; Mesquita et al. 2016; Rosa et al. 2016; Matos et al. 2018). Genotype 4 includes strains from sporadic human cases in Asia and more recently in Europe (Meng 2010; Hakze-van der Honing et al. 2011; Colson et al. 2016; Jeong et al. 2017; Wu et al. 2017a, b). Recently, a novel HEV genotype 7 was isolated from a liver transplant recipient with chronic HEV infection in the Middle East (Lee et al. 2016). Hepatitis A and hepatitis E viruses are the two main hepatitis etiological factors causing enterically transmitted infections in developing countries and among indigenous people in developed countries (Caruso et al. 2016). The mortality rate of HEV, which is usually low (1%–2%) in the general population, can be as high as 10%–25% in pregnant women, and over 75% of liver disease patients lose their livers as a result of HEV infection (Marano et al. 2015; Bazerbachi et al. 2016). A World Health Organization (WHO) report estimated that 20 million people are infected with HEV worldwide, 3.3 million of whom are symptomatic, and that ~ 56, 600 people die each year from HEV-related liver failure (WHO 2016). The possibility that it is a zoonotic disease is supported by a large body of evidences, including prevalence reports of HEV RNA detected in pigs, wild boars, rodents, and deer (Kamar et al. 2012; Song et al. 2014). From the human perspective, current evidence supports a zoonotic origin of HEV infections, with a statistically significant association identified between occupational exposure to pigs and anti-HEV immunoglobulin G (IgG) levels (Wilhelm et al. 2014). Moreover, genotyping a section of open reading frame 2 (ORF2) has shown both homology and sequence identities ranging from 70% to 100% between human- and animal-derived isolates (Wilhelm et al. 2011). With the exception of sporadic cases of HEV infection, which are predominantly caused by HEV genotype 4, large outbreaks rarely occur in China. However, the prevalence of HEV infection is high in China and it is suspected to be zoonotically transmitted from wild boars, rabbits, and domestic pigs, and pigs are thought to be a major repository of genotypes 3 and 4 (Liu et al. 2012; Dai et al. 2013; Wang and Wang 2016; Wu et al. 2017a, b). A serological survey showed that HEV infection in pigs is very common in different regions of China, with positivity rates ranging from 30% to 100% (Ding et al. 2018; Gong et al. 2018; Shuai et al. 2009; Huang et al. 2012; Zhang et al. 2010). In fact, a high seroprevalence of HEV among swine farmers and swine veterinarians has been reported in previous studies (Kang et al. 2016; Bansal et al. 2017; Lange et al. 2017; Mughini-Gras et al. 2017; Teixeira et al. 2017; Ukuli and Mugimba 2017). Residence in areas with high rates of swine contact and sharing drinking-water resources with swine have been significantly associated with HEV infection, and zoonotic spread is a major mode of HEV genotypes 4 transmission from pigs to humans in eastern China (Khuroo and Khuroo 2016).
In townships and rural areas of central China, the major pork production and the local meat economy depend on a swine breed, which is defined by the local Food and Drug Administration as an autochthonous small-sized breed of Landrace and Large White pigs. In the previous study, we investigated the acute HEV infections in commercial pigs in Anlu city of Hubei Province of China and found a high anti-HEV IgM prevalence of 46.7% in towns and an average anti-HEV prevalence of 37.9% (Zhang et al. 2016, 2017). Following this, we used seroprevalence and HEV RNA detection to assess the presence of HEV and its genotypes in the general population, swine workers, and swine in Anlu city of Hubei Province of China.
-
Anlu city of Hubei province (31°0.250'N, 113°0.690'E) is a high-density pig-farming area in central China. Sera were randomly collected from 1505 human blood samples (1232 from the general population and 273 from occupationally swine-exposed workers) during February to August in 2016. The specific sampling locations are shown in Supplementary Figure S1. The 273 swine workers included 137 pig farmers, 104 pork retailers, 24 slaughterers, and 8 veterinarians. At the time of the study, all the participants were healthy and showed no signs or symptoms of hepatitis within the 6 months preceding the survey. To assess the human exposure risk, we collected data on the demographic characteristics, employment status, and liver disease history of all subjects using a questionnaire that we designed, after conducting advanced reliability and validity analyses. Informed consent was obtained from all participants.
Forty-seven pig-fattening farms from 15 villages and towns were sampled over a 7-months period (from February to August) in 2016 (Supplementary Figure S1). Blood samples were taken from the jugular veins of 276 pigs. The two blood samples were collected at the same time from each pig and used for HEV-Ag or HEV RNA detection. The samples were centrifuged at 1000 ×g for 20 min to separate the sera, which were then stored at - 80 ℃ before analysis.
-
Acute HEV was determined by the presence of anti-HEV IgM, and previous viral exposure was confirmed by antiHEV IgG reactivity without anti-HEV IgM. The human serum samples were screened for HEV IgM and HEV IgG using anti-HEV IgM/IgG ELISA kit (Wantai Biopharmaceutical Co., Ltd, Beijing, China), according to the manufacturer's instructions. The swine serum samples were screened for HEV ORF2 antigen (HEV-Ag) using an HEVAg ELISA kit (Shanghai Meilian Biological Scientific Co. Ltd, Shanghai, China), according to the manufacturer's instructions.
-
Total RNA was extracted from 140 μL of serum collected from the human and swine, using the QIAamp Viral RNA Mini Kit (Qiagen, Germantown, MD, USA), according to the manufacturer's instructions, and then resuspended in 30 lL of DNase/RNase-free water. The total RNA was used as the template for reverse transcription (RT)-PCR with the PrimeScript First-Strand cDNA Synthesis Kit (TaKaRa Bio Inc., Kusatsu, Shiga, Japan). The 5' region of ORF2 provides a phylogenetic signal comparable to a full-genome analysis and was therefore used for a phylogenetic analysis (Lu et al. 2006). After nested RT-PCR was performed, the amplicons were sequenced. In brief, the HEV ORF2 domain sequences were amplified using RT products as the templates. The two pairs of primer sets were: 5'-CCCTTATCCTGCTGAGCATTCTC-3' (F) and 5'-AAY TATGCMCAGTACCGGGTTG-3' (R) (first primer set), and 5'-GTYATGYTYTGCATACATGGCT-3' (F) and 5'-AGCCGACGAAATYAATTCTGTC-3' (R) (second primer set), with product lengths 731 bp and 438 bp, respectively. The PCRs were run on an EppendorfTM MastercyclerTM pro PCR System (Thermo Fisher Scientific, Inc., Hercules, CA, USA) under the following threestep cycling conditions: one initial template denaturation step at 95 ℃ for 2 min; 35 cycles of denaturation at 95 ℃ for 15 s, annealing at 50 ℃ for 30 s, and elongation at 72 ℃ for 50 s; and a final elongation step at 72 ℃ for 7 min. The second primer set was used during the second round of PCR, using the same procedure. The amplification products were analyzed on a 1% agarose gel, and then purified with a PCR clean-up system (QIAquick PCR Purification Kit, cat no./ID: 28104, Qiagen). After purification, the amplicons were sequenced at Sangon Biotech Company (Shanghai, China).
-
Forth-three HEV strains with different genotypes were selected as reference sequences for constructing phylogenetic trees. A phylogenetic analysis was performed with the MEGA version 6 (Tamura et al. 2013) software. We constructed phylogenetic trees with the neighbor-joining method and evaluated them with the interior branch test method. The Kimura model was then used to calculate the genetic distances. The statistical robustness and reliability of the branching order were confirmed with a bootstrap analysis of 1000 resampled data points. The similarity and divergence ratios of the strains were calculated with the Lasergene software package MegAlign (DNASTAR, Madison, WI, USA).
-
We descriptively estimated the seroprevalence of HEV in humans after grouping them by age, sex, and work experience. The rates were compared with either the χ2 or Fisher's exact test. We estimated the odds ratios (ORs) and Wald's 95% confidence intervals (CIs) for the conditional logistic regression models to identify significant risk factors in the occupationally swine-exposed group. We used Wald's χ2 to test the significance of each item in the model. Statistical significance was set at P < 0.05.
Study Design for Human and Swine Sampling
Serological Analyses
HEV RNA Detection and Isolation of the Partial ORF2 Region
Phylogenetic Analysis
Statistical Analyses
-
Among the 1505 human serum samples tested, 455 subjects (30.23%) from the study cohort were positive for anti-HEV IgG. Out of these, 30 (1.99%) were also positive for antiHEV IgM and 7 (0.47%) were positive for HEV RNA. Further analysis showed that the prevalence of anti-HEV IgG in the 273 workers with occupational swine contact was significantly higher than in the 1232 representatives from the general population (χ2 characteristic of independence [χ2] for this distribution = 51.908, with degrees of freedom [df] = 5, P < 0.001; Table 1). No participants had any history of travel abroad. Among the 276 swine serum samples, we found a prevalence of 6.52% (18/276) for HEV-Ag and 2.54% (7/276) for HEV RNA.
Variables No. detected Positive numbers Positive rate (%) Chi square test Crowd χ2 = 51.908 General population (GP) 1232 323 26.22 P = 0.00 Workers occupationally contact with swine 273 132 48.35 Total 1505 455 30.23 Gender in GP χ2 = 10.874 Male 582 178 30.58 P = 0.01 Female 650 145 22.31 Total 1232 323 26.22 Age grouping in GP (years) χ2 = 160.925 0–5 165 3 1.82 P = 0.00 6–10 89 5 5.62 11–15 61 4 6.56 16–20 96 9 9.38 21–30 104 24 23.08 31–40 131 43 32.82 41–50 133 49 36.84 51–60 113 48 42.48 61–70 99 44 44.44 71–80 181 69 38.12 81–90 60 25 41.67 Total 1232 323 26.22 Occupational grouping in GPs χ2 = 155.871 Preschool children 181 5 2.76 P = 0.00 Students 219 15 6.85 Peasants 129 60 46.51 Blue-collar workers 112 42 37.5 Managerial staffs 102 39 38.24 Medical staffs 84 18 21.43 Clerks in public place 121 45 37.19 Others 284 99 34.86 Total 1232 323 26.22 Table 1. The positive rates of anti-HEV IgG among recruited cases.
-
We divided the general population into 11 age subgroups, ranging from 0 to 90 years old. Table 1 shows that 323 (26.22%) out of the 1232 people from the general population had detectable anti-HEV IgG levels. Surprisingly, the prevalence of anti-HEV IgG ranged from its lowest value (1.82%) in the 0 to 5-years age group, up to its highest value (44.44%) in the 61 to 70-years age group, and the age-specific differences in prevalence were statistically significant (χ2 = 160.925, P < 0.001), with a linear distribution. The male:female ratio for anti-HEV IgG was 1.22:1, which indicated a significant difference by sex (χ2 = 10.874, P = 0.01), insofar as the prevalence was higher in men (30.58%, 178/582) than in women (22.31%, 145/650). Among the general population, we expected occupation to be a key differentiating factor for the antiHEV IgG positivity. In fact, we found a significantly higher occurrence of HEV infection in peasants (46.51%) than in other groups, whereas the prevalence was lowest in preschool children (2.76%). Our result showed high prevalence of HEV among blue-collar workers (37.50%), managerial staff (38.24%), and clerks in the public (37.19%) compared with the other occupational groups. Retirees and other people not belonging to specific occupational groups were classified into the "others" group, and the prevalence in this group was high, at 34.86% (Table 1).
-
Anti-HEV IgG is a standard marker of an immune response to previous HEV infection. We used a multivariable model to identify the factors that predicted anti-HEV IgG positivity among the 273 workers with occupational swine contact. We included age, working years, sex, education level, occupational pattern, and working hours per day in the model. Overall, younger workers were significantly less likely to show evidence of previous HEV infection (OR = 2.067, χ2 = 14.649, P < 0.001; Table 2), and people > 40 years old had a higher HEV-infection risk and were more likely to test positive for anti-HEV IgG than people < 30 years old. A similar trend was also detected in working years (OR = 6.0, χ2 = 11.903, P = 0.018), and more working years was associated with an increased risk of HEV. The statistical analysis showed no significant differences in the sex (P = 0.491), education level (P = 0.321), occupation (P = 0.163), or working hours per day (P = 0.331) among workers who had swine contact.
Factors Variables β Wald c2 P value OR (95% CI) Chi square test Gender 1 = male, 0 = female 0.15 0.474 0.491 1.162 (0.759–1.779) Age group (years) 24.699 0 2.067 χ2 = 14.649 1≦30 years -1.371 12.969 0 0.254 (0.120–0.535) P = 0.002 2 = 31–40 years -0.824 6.551 0.01 0.439 (0.234–0.825) 3 = 41–50 years -1.044 19.379 0 0.352 (0.221–0.560) Years of Working 23.243 0 6 χ2 = 11.903 1 = 0–5 years, -3.007 13.445 0 0.049 (0.010–0.247) P = 0.018 2 = 6–10 years, -2.772 11.946 0.001 0.063 (0.013–0.301) 3 = 11–15 years, -2.986 13.313 0 0.050 (0.010–0.251) 4 = 16–20 years, -1.576 2.881 0.09 0.207 (0.033–1.276) Educational background 3.5 0.321 0.923 χ2 = 1.749 1 = Primary school -0.811 1.987 0.159 0.444 (0.144–1.373) P = 0.417 2 = Junior school -0.483 0.769 0.381 0.617 (0.210–1.815) 3 = Senior school -0.826 2.067 0.151 0.438 (0.142–1.350) Job pattern 1.95 0.163 1.2 χ2 = 5.637 1 = pig farmers 1.713 7.134 0.068 5.545 (1.011–30.424) P = 0.060 2 = Slaughters 2.396 3.89 0.049 10.984 (1.650–73.130) 3 = Pork retailers 2.303 6.139 0.013 10.009 (1.533–65.349) Working hours per day 2.214 0.331 1.506 χ2 = 8.077 1≦6 h -0.008 0 0.985 1.008 (0.452–2.247) P = 0.018 2 = 7–9 h -0.398 1.012 0.315 0.670 (0.307–1.462) Table 2. Analysis for influent factors of the positive rates of anti-HEV IgG among workers occupationally contacted with swine using logistic regression model.
-
There was no significant difference in anti-HEV IgM prevalence between males and females (1.83% [14/767] and 2.17% [16/738], respectively). There was also no any significant age-related (χ2 = 0.268, P = 8.793) or occupation-related differences in anti-HEV IgM prevalence detected (Table 3). The epidemiological questionnaire showed that two out of the 30 patients with anti-HEV IgM were also diagnosed with liver dysfunction; one was female with no history of pig exposure and the other was a male swine worker. The female patient was coinfected with HBV and HCV, and presented with chronic liver disease without jaundice. Her alanine aminotransferase (ALT), aspartate aminotransferase (AST), total bilirubin (TBIL), and direct bilirubin (DBIL) levels were 92.9 U/L, 108.9 U/L, 13.63 μmol/L, and 4.6 μmol/L respectively. The ALT, AST, TBIL and DBIL levels of the male patient were 462.5 U/L, 138.4 U/L, 155.37 μmol/L and 115.73 μmol/L, respectively, and were accompanied by evident clinical jaundice. Only two of the 30 people with anti-HEVantibody-positive sera had abnormal liver function, which suggests that HEV-infected people often have asymptomatic liver disease.
Variables No. detected Positive numbers Positive rate (%) Chi square test Gender χ2 = 0.634 Male 767 14 1.83 P = 0.226 Female 738 16 2.17 Age grouping (years) χ2 = 0.268 0–10 254 0 0 P = 8.793 11–20 161 1 0.62 21–30 132 0 0 31–40 174 7 4.02 41–50 246 7 2.85 51–60 189 4 2.12 61–70 107 4 3.74 71–80 182 5 2.75 81–90 60 2 3.33 Occupation grouping χ2 = 76.158 Preschool children 181 0 0 P = 0.000 Students 219 1 0.46 Peasants 129 10 7.75 Blue-collar workers 112 0 0 Managerial staffs 102 0 0 Medical staffs 84 0 0 Clerks in public place 121 11 9.09 Others 284 0 0 Pig farmers 137 3 2.19 Pork retailers 104 5 4.81 Slaughterers 24 0 0 Veterinarians 8 0 0 Table 3. The positive rates of anti-HEV IgM using Chi square test among general population and workers occupationally contacted with swine.
-
A sequence of 100% homology in the same species is considered to be the same sequence. There were 4 different sequences after comparing and analyzing the homology of 7 swine HEV ORF2 sequences. After the homology analysis of 7 human HEV ORF2 sequences, 4 different sequences were found. Homology and phylogenetic analyses were performed on the partial ORF2 sequences from pig samples (GenBank nos KX463414–KX463417) and human samples (GenBank nos KX463410–KX463413). The human and swine HEV isolates shared 97.04%–100% sequence homology, and ORF2 from a human sample (KX463411) shared 100% sequence identity with ORF2 from a swine sample (KX463417). The four human HEV isolates shared 97.04%–99.34% sequence homology and the swine isolates shared 98.68%–99.67% sequence homology. The eight HEV isolates also shared 75.33%–79.28%, 77.96%–78.95%, 75.33%–78.62%, and 82.89%–94.08% identity with GenBank reference sequences of HEV genotypes 1–4, respectively. Homology and phylogenetic analyses confirmed that the eight novel HEV sequences detected in this study were subtype 4d, and shared 96.05%–97.37% nucleotide sequence identity with the most closely matched strain (GenBank no. FJ46 1765), which was isolated from a person in Hubei Province (Fig. 1), and 89.14%–90.46% identity with another strain (GenBank no. FJ46 1762) that was isolated from a swine in Hubei Province. Further analysis of the ORF2 amplicons revealed higher sequence homologies (96.05%–97.04% and 95.72%–96.38% identity) with genotype 4d from Shandong (KF176370) and Beijing (KP196807) HEV swine strains, respectively.
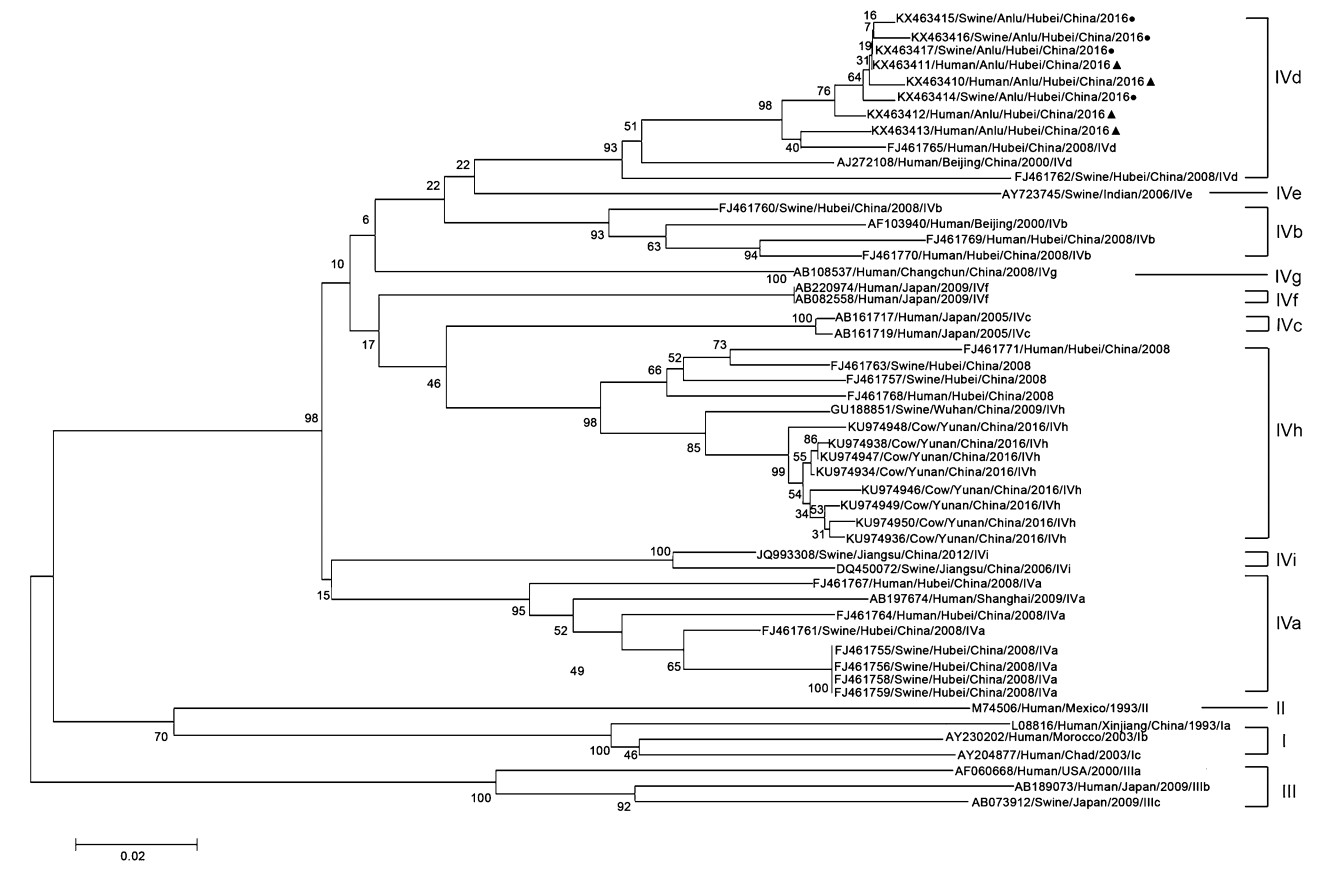
Figure 1. Phylogenetic analysis of HEV isolates from human populations and swine herds. The phylogenetic tree was constructed by the neighbor-joining method and evaluated by interior branch test, based on partial nucleotide sequence of the ORF2 region. Forty-three known representative strains published in GenBank were used as references. The human or swine's isolates were signed with filled triangle or filled circle, respectively.
Demographic Characteristics of HEV Seroprevalence in Individuals
Prevalence of Anti-HEV IgG among the General Population according to Sex, Age, and Current Occupation
Multivariable Conditional Logistic Regression Analysis of Risk Factors for HEV Infection among Occupationally Swine-Exposed Workers
Analysis of Epidemic and Clinical Characteristics of 30 Anti-HEV-IgM-Positive Patients
Phylogenetic Analysis of HEV ORF2 RNA Sequence
-
In Asia and Africa, HEV is generally considered an endemic disease that causes acute self-limiting hepatitis in young people, with the exception of pregnant women, in whom fatality rates are usually 10%–25% or greater (Kamar et al. 2012). In Spain, the HEV rate has remained stable at 12%–17%. In European Union and European Economic Area countries, HEV infections are predominantly indigenous and are caused by HEV genotype 3, the commonest viral genotype in the human and animal reservoirs in Europe (Mesquita et al. 2016; Adlhoch et al. 2016; Rosa et al. 2016; Matos et al. 2018). Several crosssectional studies in Norway, Canada, and other countries have also shown an increased HEV-infection risk in people with direct exposure to pigs or pork products (Walachowski et al. 2014; Kang et al. 2016; Bansal et al. 2017; Lange et al. 2017; Mykytczuk et al. 2017; Mughini-Gras et al. 2017; Teixeira et al. 2017; Ukuli and Mugimba 2017). In Europe, anti-HEV antibody prevalence among the general population was 1.3%–52%, whereas a high seroprevalence of HEV IgG (13%–51.1%) was detected among swine farmers (Lapa et al. 2015). Surprisingly, antiHEV IgG prevalence was very low (5.6%) among swine farmers in Croatia (Vilibic-Cavlek et al. 2016). In a previous study, we investigated acute HEV infection in commercial pigs in Anlu city of Hubei Province, and found a high anti-HEV IgM prevalence of 46.7% in towns and an average anti-HEV-antibody-positive prevalence of 37.9% (Zhang et al. 2016, 2017). Our results indicate that HEV is prevalent in pigs in Anlu city of Hubei Province and that the seroprevalence among workers who are occupationally exposed to pigs (48.35%) is almost twice that of the general population not exposed to pigs (26.22%). This indicates an association between direct swine contact and a high risk of HEV infection.
In addition to global variations in the epidemiology of HEV, the seroprevalence of HEV within a single country can vary widely. Consistent with previous findings, we found that anti-HEV IgG prevalence increased with age, with people > 50 years old more likely to be infected with HEV than people < 50 years old (Kamar et al. 2014; Taherkhani and Farshadpour 2016; Lange et al. 2017). We also reported that the overall prevalence of anti-HEV IgG in people < 20 years old was only 5.11% (21/411), which is below the 14.93% reported in northern China or even the 10% estimated in the Netherlands (Meng et al. 2015; van et al. 2017). As expected, the mean prevalence of antiHEV IgG among people aged ≥ 50 years exceeded 40% in the general population of Anlu city, which is higher than the 33% reported in the Netherlands among similarly aged people. This further supports the conclusion that viral exposure has a cumulative age-dependent lifetime pattern that correlates with decreasing immunity (van et al. 2017). This result also indicates that previous infections reflect an age-cohort effect. In the majority of studies, HEV seroprevalence has been higher in males than in females and HEV antibody positivity has also been higher in male blood donors and males in the general population (Mohebbi et al. 2012; Fearon et al. 2017; van Gageldonk-Lafeber et al. 2017). We also found a significant sex difference in exposure rate (χ2 = 10.874, P = 0.01), which may be attributable to the higher frequency with which men in China participate in business entertainment, including eating dinner outside the home, compared with women. We also found that HEV seroprevalence (antiHEV IgG) differed significantly by occupation among the general population, insofar as only 2.76% of preschool children were positive for anti-HEV IgG, whereas 46.51% of peasants in the general population were positive. This finding may indirectly support the prediction that poor sanitation increases the risk of HEV infection. Age and working years were the main factors positively associated with HEV seropositivity among workers with occupational swine contact (χ2 = 14.649, P = 0.002 and χ2 = 11.903, P = 0.018, respectively), which is consistent with a previous study (Song et al. 2014).
Several studies have found that genotype 4 is not only prevalent in developed countries, but also distributed across China, Vietnam, and India, where subtypes 4a–d have been identified (Tian et al. 2015; Bansal et al. 2017; Jeong et al. 2017; Mykytczuk et al. 2017). Subtype 4d is mainly distributed in China and Italy, among both humans and swine (Garbuglia et al. 2013; Monne et al. 2015). In this study, we also found that HEV genotype 4 was the main genotype, and a phylogenetic analysis confirmed that this subtype was 4d. Using nested RT–PCR, followed by amplicon sequencing, we amplified 14 HEV sequences, seven sequences from human populations and another seven from swine herds. A homology analysis identified four novel human HEV sequences and four novel swine HEV sequences. We detected sequence identity of 100% between one swine HEV sequence (KX463417) and one human HEV sequence (KX463411); in total, the four human HEV sequences and four swine HEV sequences shared 97.04%–100% homology. Therefore, we speculate that HEV genotype 4d is epidemic in this specific local region, and that swine may be one of the sources of the HEV genotype 4d infections in the local human population. In fact, one older HEV-infected man was also a pig farmer. Interestingly, the 304 bp HEV ORF2 sequences that we sequenced shared only 89.14%–90.46% homology with another previously identified sequence (FJ46 1762) from Hubei, which further supports the argument that epidemics of different HEV subtypes have occurred in the local region, rather than a widespread epidemic of a single genotype. By contrast, genotype 4d identified in this study shares greatest sequence homology with Shandong (KF176370) and Beijing (KP196807) strains, which suggests that this genotype 4d strain was imported to the city of Anlu from other provinces or cities.
This study had several limitations. First, we only recruited individuals from Anlu city of Hubei province, so our estimates of HEV seroprevalence cannot be generalized across central China. Further studies should be performed in other cities with pig-breeding activities similar to those in Anlu. Second, pig-farm workers who were exposed to pigs in the swine age group with the highest viral shedding rate also had the highest risk of HEV infection (Wilhelm et al. 2016). However, we did not subdivide this group from the 273 members of the occupationally swine-exposed group, so did not control for this effect. Third, we did not collect data on other potential HEV-infection-related activities, such as alcohol use, diet, or pork consumption.
-
The authors would like to thank the Anlu animal husbandry and veterinary bureau for these supporting information on pig density at Anlu city in the central China in 2016. This work was partly supported by General Projects of Health and Family Planning Commission of Hubei Province of China no. WJ2017M174, and WJ2017M240 and Occupational Hazard and Identification Control of Hubei Provincial Key Laboratory Open Fund, no. OCHI2017G02.
-
YS, JZ, LZ and HW designed the study. YS, YC, SZ and QW performed the experiments. YS, SZ, CZ and ZZ analyzed the data. HC, YS and LZ drafted the manuscript. All authors read and approved the final manuscript.
-
The authors declare that they have no conflict of interest.
-
All institutional and national guidelines for the care and use of animals were followed. Additional informed consent was obtained from all patients for which identifying information is included in this article.

 DownLoad:
DownLoad: