











-
Rabies is one of the better known encephalitides, caused by the rabies virus of the genus Lyssavirus in the family of Rhabdoviridae (George M B, 2007). About 55000 human rabies cases are reported each year worldwide with ≥ 90% caused by dog (Canis lupus familiaris) bites (Zhang Y Z, 2006). In countries where dog rabies has been controlled due to active vaccination campaigns and surveillance, dog rabies is no longer prevalent, and human cases in these countries or regions are usually caused by wildlife reservoirs (Rupprecht C E, 2006). Foxes, raccoons, skunks, jackals, mongooses, and bats have been successively reported to be responsible for human deaths in these regions (Hanlon C A, 2007). Nevertheless, in China, dog rabies is still considered to serve as the main source of infection for human rabies (Hu R L, 2008) and in the total number of 2000-3000 human rabies cases reported annually at the national level, ≥ 95% were dog-associated rabies, according to retrospective epidemiology investigation. Cat, mongoose and other wildlife-associated human rabies in China have been generally considered to be infrequent and isolated incidents result from spillover from rabid dogs without epidemiological significance.
In an independent survey investigating Chinese Ferret Badger-associated human rabies, we found that ferret badger rabies cases in some counties in Zhejiang and Jiangxi provinces were more prevalent than dog-associated in the same regions (Wang Z, 2006; Zhang S F, 2009). For example, from 2002-2004 in Hangzhou district, Zhejiang province, 29 out of 42 human rabies cases were badger-associated. After the successive outbreaks of badger-associated human rabies, the local center for disease prevention and control (CDC) advised residents to be careful to avoid contact with these kinds of animals, and once bitten, a rabies Post-Exposure Prophylaxis (PEP) regimen should be started immediately. However, these recommendations have not been made in other regions such as in Jiangxi province. With the increase of badger-associated human rabies, we hypothesize that the rabies transmission in ferret badgers has formed an independent cycle. However, evidence about whether an independent transmission cycle exists in badger populations or the badger rabies was just a spillover from dogs is lacking. To investigate this hypothesis, from 2007-2010, we performed an extensive epidemiology and molecular epidemiology study in Wuyuan, Leping and Jingdezhen counties, Jiangxi province. 15 rabies viruses from ferret badgers in these counties in Jiangxi province were isolated as part of this surveillance program, combined with other publicly available isolates and investigated from an epidemiological standpoint.
HTML
-
The information on human rabies cases were obtained from the Jiangxi provincial CDC, which was obtained by retrospective investigating the epidemiology for human cases. For each human case suspected to be associated with rabies, detailed information, including bite history, is collected from the subject after they were sent to hospital as described elsewhere (Yu J N, 2012).
-
Dead ferret badger brain samples were collected from Wuyuan, Leping, Fuzhou and Jingdezhang counties in Jiangxi province, and Lin'an, Chun'an, Quzhou, Taizhou counties in Zhejiang province between January 2007 to December 2009. Farmers or residents were requested to send the dead and undecayed animals to the collecting sites of the animals.
The brains of ferret badgers were detected for rabies virus infection by direct fluorescence assay (DFA) as described elsewhere (Beauregard M, 1965). Briefly, smears of brain specimens were prepared on glass slides, and then fixed in 80% ice-cold acetone, stained with FITC-conjugated anti-nucleoprotein monoclonal antibody (in-house preparation). The stained slides were observed under a fluorescence microscope (Wang W J, 2012).
-
Once a ferret badger brain was confirmed to contain rabies antigen, it was homogenized in 10 volumes (w/v) of phosphate-buffered saline (PBS, pH 7.2), and 20-30 µL of the suspension was injected intracerebrally into a brood of 1-day-old suckling mice (Animal Center, Institute of Changchun Biological Products, Changchun) respectively. The mice were observed to 28 days post-injection for clinical signs indicative of rabies infection. Once clinical signs appeared, the mice were killed humanely and the antigens in the brains were confirmed by DFA.
-
The N, M and G genes of the rabies virus isolates were amplified from the ferret badger brain samples or the infected mouse brain by reverse transcription-polymerase chain reaction (RT-PCR) according to the protocol described elsewhere (Nadin-Davis S A, 1994) using validated primers (Zhang S F, 2010), the primers positions were according to reference strain FJ009, GenBank No. FJ866836.1.
The RT-PCR assays of the infected ferret badger or mouse brain was repeated three times. Sequencing of the amplicons was carried out by TaKaRa Corp. Ltd, Dalian, China. The obtained sequences were compared with the sequences of rabies virus isolates from other ferret badgers and dogs isolates published in GenBank. The Neighbor Joining method was employed for phylogenetic analysis in software MEGA 4 (MegAlign, DNASTAR v.7.1.0). Bootstrap values were calculated from 1000 repeats.
-
The amino acid sequences of the nucleoprotein, glycoprotein and matrix protein were aligned and compared among all the isolates from ferret badgers, together with some representative published isolates from dogs, by the software MEGA 4 (MegAlign, DNASTAR).
The epidemiological survey
Specimens and direct fluorescence assay (DFA)
Isolation of the rabies virus and confirming diagnosis
Amplification and sequencing of the 3 most conserved genes of the isolates
Amino acid sequence analysis of the 3 most conserved genes of the isolates
-
From 2007-2009, 6 people (3 in 2007, 2 in 2008 and 1 in 2009) died of rabies in Wuyuan county, Jiangxi province, while there were no dog-associated human rabies cases anywhere in this county. Of these six cases, 3 were hunters and 3 were farmers. The contact between the hunters and badgers were frequent and direct as they manage the animals for sale almost every day, and they were not generally concerned with injuries when bitten. This is highlighted by three cases. The first case was a female farm worker who found a sick badger when walking along the road, when she tried to catch the badger on its tail, the badger bit the hand of the woman; in the second case a man found 3 badgers playing with each other in the field, and was bitten when he tried to separate them in order to catch one of them. The third one was bitten in his house when he got up at midnight; a sick badger had entered his house and bit him on the foot when he stepped on it. None of the cases sought post-exposure prophylaxis (PEP). All 6 people died, 5 died 2-3 months after the bite, and 1 died 1 year after the bite.
-
From the beginning of 2007 to the end of 2009, a total of 178 ferret badger specimens were collected from Jiangxi and Zhejiang province. After DFA assay, 15 (8.9%) of them were found positive. Virus isolates were obtained from all the positive samples (Table 1).

Table 1. Background information for isolates collected from ferret badger brain specimens as part of the rabies wildlife surveillance program
-
Both the rabies virus positive ferret badger brains and the infected mouse brains were used for gene amplification and sequencing. The 15 sequenced isolates obtained in this study in Jiangxi and Zhejiang provinces were aligned, compared and a phylogenetic tree was constructed (Fig. 1). The tree contained three major clades (Clades 1 to 3) with strong bootstrap support. 14 of the 15 isolates formed a single sub lineage within Clade 2. They shared 94.7%-100% nucleoprotein homology with one another, but only 87.8%-95.7% homology with other dog origin isolates in Clade 2, 88.0%-90.2% homology with isolates in Clade 1, and 85%-87.9% homology with the dog isolates comprising Clade 3. There one ferret badger exception, JX09-17, collected in this study which didn't place in the main ferret badger clade was isolated from Fuzhou, a county of Jiangxi province and far from where the 14 isolates were isolated (Zhang S F, 2010). The nucleotide sequence of both nucleoprotein and glycoprotein shared higher homology with the dog isolates, but showed some differences in amino acid sequence for the glycoprotein and was considered a variant of dog rabies virus origin. Up to now, no dog rabies virus isolate has been reported to be closely associated with these isolates (Fig. 1).
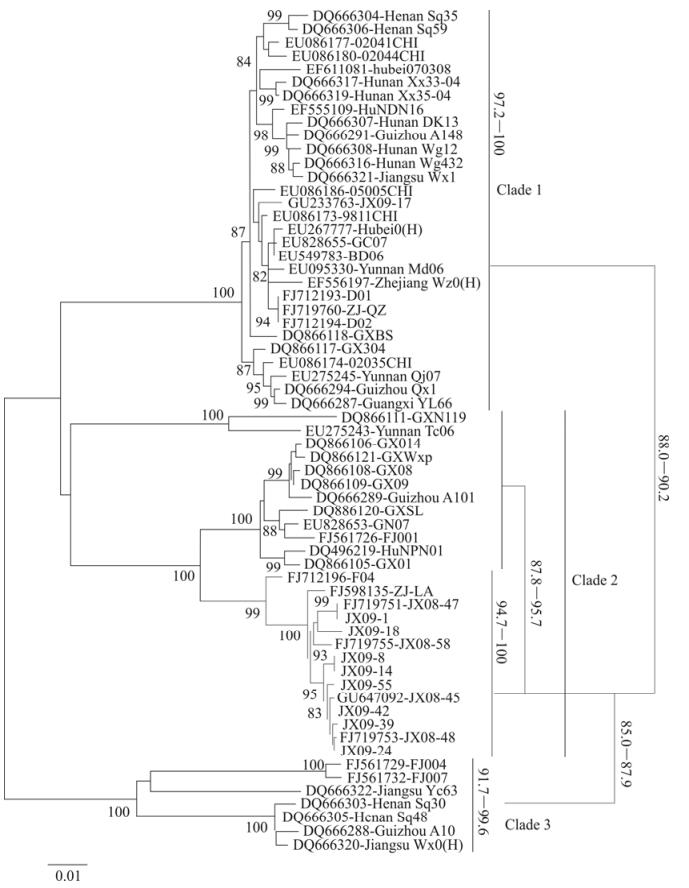
Figure 1. Phylogenetic tree of nucleoprotein of ferret badgers rabies virus isolates. 14 of all the 15 ferret badgers rabies virus isolates (marked in red) are clustered into a single lineage, The homologies are 85.0% to 95.7% between ferret badger and dog isolates compared to 94.7% to 100% amongst ferret badger isolates.
It can also be inferred from Fig. 1 that the rabies viruses maintained in ferret badger populations may share some common ancestor with dog isolates in Clade 2, but have experienced long term of adaption in ferret badgers (based on the branch lengths and observed nucleotide mutations).
-
Comparison of the amino acids among the 14 isolates and with other dog isolates showed that the amino acid sequences of these 14 isolates from ferret badgers are highly conserved, ranging from 99%-100%. However, they shared 96.7%-98.8% homology with the isolates in Clade 2; 96.4%-98.3% with Clade 1, which is the emerging lineage in the current rabies epidemic in China (Yu J N, 2012); 94.5%-96.7% with the isolates in Clade 3. The high homology between the isolates from ferret badgers and the other isolates in Clade 2 also indicates a shared common ancestor at some earlier time. There are characteristic amino acid mutations in the ferret badgers amino acid sequences at sites 181 and 374 in the nucleoprotein, at sites 201, 408 and 467 in the glycoprotein, and at site 70 in the matrix protein as shown in Table 2. These results also reflect the long term adaptation in ferret badgers. Whether these typical amino acids represent interspecies transmission requires further investigation. It should be noted that all the amino acids at glycosylation sites and the most recognized major antigenic sites keep unchanged.

Table 2. The characteristic amino acid signatures identified in the major structural proteins of rabies virus isolates from ferret badgers
Human rabies cases in study area and compared to other rabies incident regions in China
Rabies virus positive specimens and isolation of viruses
Nucleotide epidemiology of rabies isolates shows the independent lineage
Unique signature amino acid mutations are identified in isolates from ferret badgers
-
Since 1994, badger-associated clinical human rabies has been reported in different counties in Zhejiang and Anhui provinces. However, due to reduced rabies surveillance in China in previous years, studies on rabies and rabies virus was not extensive and no rabies virus isolates from ferret badgers were obtained. Even in 2002 to 2004, when dog associated-human rabies in China became increasingly serious, ferret badger-associated human rabies were limited to Hangzhou and Zhejiang province, and no funding was available to support further investigation. Therefore, rabies virus of ferret badger origin was not isolated until 2007 and there is a paucity of information about ferret badger rabies, both from a biological and epidemiological perspective. In recent years, National Natural Science Foundation of China (NSFC) and national "973" and "863" projects have supported some basic researches on wildlife rabies. As a consequence we have been able to collect the specimens from badgers since 2007 in Jiangxi and Zhejiang provinces.
As there is limited public education on rabies in China there is correspondingly limited awareness of the potential threat from the disease. Consequently, people in rural areas rarely send dead animals to animal quarantine departments for analysis and collection mainly depends on our research group and some farmers and hunters who are engaged in animal trade. However, the area for specimen collection in this study is broad enough to incorporate different counties and different provinces to achieve some degree of representativeness.
When ferret badger-associated human cases were firstly recognized as a public health problem in Hangzhou and Zhejiang province, government propaganda to raise awareness of the threat from ferret badger bites was extensive and consequently ferret badger-associated human rabies cases have fallen. However, no such public information messages were disseminated to the residents in the Wuyuan county, and rabies in ferret badgers is new to both residents and medical doctors in the CDC in this county, thus the greatest number of cases have occurred in this area for the previous 3 years. As the disease prevention and control system in China is becoming more established, it is hoped that these events will not be repeated the in human population in the future.
With the economical development of China in the last twenty years, the lands and the hills become fertile, and the vegetative cover and the number and density of wildlife has substantially increased. Ferret badgers are one of the major wildlife distributed widely all over China, but mainly reside in mountain and lake areas. Ferret badger-associated human rabies was firstly reported in 1994-1995 in Huzhou city, Zhejiang (Shi B N, 1997). Afterwards, small scale outbreaks continued to occur in human populations between 2002-2004 in both Huzhou and Hangzhou, Zhejiang (Li Y X, 2004). We recognized the presence of a rabies epidemic in wildlife 2005 and were able to isolate the first rabies virus strain from a rabid ferret badger in 2008 (Zhang S F, 2009). Apparently, it was related to dog isolate but phylogenetic analysis placed it in an independent lineage of Clade 2, apparently distant from isolates from dogs. With the increasing of the number of isolates from ferret badgers in these 3 years from a wide range of areas, we found these isolates still aggregates together (with one exception derived from Fuzhou), and seemed to be a definite independent lineage of others, distinct from dog isolates. This is further supported by the identification of characteristic amino acid mutations in the nucleoprotein, glycoprotein and matrix sequences that are not present in any dog isolates. As these ferret badger samples were collected in different provinces at different times, this indicates that the virus infecting ferret badgers is not a recent spillover and the two lineages have circulated independently for many years. This further suggests the virus has undergone a host adaption in the ferret badgers. However, the conservation of glycosylation sites and the majority of major antigenic sites underlie their importance for rabies in the virus life cycle and for maintaining pathogenicity or immunity.
The representative dog rabies virus isolates were selected from different areas in China and from different periods and represent different sources of rabies isolates. When we attempted to collect dog rabies or dog-associated human rabies specimens in our study, we found that in Wuyuan county, Jiangxi province, all 6 human cases reported to CDC were associated with ferret badgers, no dog-associated human rabies cases were reported in this county since 1995 and dog rabies virus from adjacent counties showed distinct differences with the virus in ferret badgers (Liu Y, 2010).
Until present, there is no effective sampling mechanism to ensure the continued surveillance of ferret badger rabies. As it is hard to compare the prevalence of rabies in dogs and ferret badgers in same region, it may be possible to distinguish between dog and rabies cases by differentiating signatures mutations in these two hosts in order to determine the transmission direction.
In conclusion, in the 3 years of our study period, 6 cases in humans in the study area have been linked to ferret badgers, highlighting the public health risks associated with rabid ferret badgers. Our data in this study showed that the rabies in Chinese ferret badgers has formed an independent transmission cycle based on their molecular phylogenetic relationship with dog rabies virus isolates in China, and represent a second rabies reservoir. Thus, it is necessary to monitor the transmission among other wildlife populations, and establish surveillance on spillover of ferret badger rabies to cats or dogs. Contact between human and these kinds of animals should be considered to carry a serious risk of exposure to rabies. Thus, oral vaccination should be initiated to halt the spread of rabies among ferret badgers and subsequent human cases.


 DownLoad:
DownLoad: