












-
Dear Editor,
Human parechoviruses (HPeVs) are members of the Picornaviridae family and consist of a non-enveloped positive-sense single-stranded RNA(+ ssRNA)(Kadambariet al. 2019; Sridhar et al. 2019). The Parechovirus genus is comprised of six species, Parechovirus A, B (formerly named Ljungan virus), C (Sebokele virus), D (ferret parechovirus), E (falcon parechovirus) and F (gecko parechovirus). Their 7.3 kb genome contains 3 regions (P1–P3), which encode a polyprotein. The P1 region encodes the structural proteins of the virus in the following order VP0, VP3 and VP1, which form the icosahedral capsid (Benschop et al. 2010).
The species of Parechovirus A infects only humans and 19 different HPeV genotypes have been identified, with HPeV-1, HPeV-2 and HPeV-3 representing the most common circulating genotypes (www.picornaviridae.com). These genotypes involved in different clinical manifestations ranging from gastroenteritis and respiratory illness to sepsis-like syndrome (Zhao et al. 2016).
HPeVs were underrecognized as a cause of meningitis and encephalitis in children, until the routine use of multiplex platforms that include HPeVs in central nervous system (CNS) pathogen detection (Kadambari et al. 2019). HPeV CNS infections are usually difficult to distinguish from other viral or bacterial infections based on clinical signs and symptoms and unnecessary use of antibiotics and antivirals is common, especially in neonates (Posnakoglou et al. 2020).
HPeVs are rapidly evolving viruses via recombination events and substitutions leading to the creation of new genotypes, which could be associated with atypical or severe clinical manifestations (Graul et al. 2017). Limited recent data have been published regarding the molecular epidemiology of HPeVs. This study aimed to investigate the molecular epidemiology and the genetic variations in the VP1 region of HPeV involved in pediatric CNS infections.
This prospective cohort study included 330 children (0–16 years) who were admitted to "Aghia Sophia" Children's Hospital with a possible clinical diagnosis of CNS infection from October 2017 to September 2020. This is the largest tertiary pediatric hospital in Greece which serves almost 40% of the metropolitan Athens area and is a reference center for Southern and Central Greece. During the 3-year study period, a total of 330 different CSF samples from children with suspected CSF infection were tested with the multiplex PCR FilmArray® ME panel and 13 (4%) bacterial and 92 (27.9%) viral infections were detected (Posnakoglou et al. 2021). HPeV was detected in 6/330 (1.8%) CSF samples, representing 6.5% (6/92) among the detected viral CNS infections.
All cases were identified in infants less than 3 months, and in this age group HPeVs represented 11.6% (6/51) of viral meningitis cases. The median age of the HPeV positive infants was 0.6 months (IQR: 0.5–1.6) and 83.3% (5/6) were male. Neonates (6/6) presented with acute febrile disease, 3/6 presented with a sepsis-like illness, 3/6 with a rash, and 1/6 with seizures (Table 1). All infants were administered antibiotics and one neonate with a severe sepsis-like illness had also received intravenous immunoglobulin. In 3/6 neonates, the empirical antibiotic treatment was discontinued after HPeV detection. All neonates with HPeV recovered without obvious sequelae.
Child Gender Age(days) Fever Rash Seizures Sepsis-like illness WBC/mm3 in CSF WBC/mm3 in Blood Neutrophils(%) Lymphocytes(%) Protein(g/dl) Glucose(mg/dl) CRP(mg/l) Hospitalization(days) Treatment(days) 1 F 45 + - - + 17 5490 75 14 32 39 11 3 2 2 M 20 + + - - 5 6880 75 16 93 94 1 9 8 3 M 15 + + - + 0 4330 47.4 30.8 86 40 4.49 9 9 4 M 60 + - - - 8 2560 24.8 52.9 62 42 18.9 3 3 5 M 15 + - + + 5 4770 38.3 31.6 60 45 5 4 4 6 M 20 + + + - 2 8540 35.1 45.9 86 48 1 6 6 The symbol " + " indicates the presence of the symptom and the symbol " -" its absence; F = female; M = male; WBC = White Blood Cell; CRP = C-reactive protein. Table 1. Demographic, clinical and laboratory data of 6 neonates who were infected by HPeV-3.
CSF was tested with conventional microbiological procedures including CSF analysis (cell counts, protein and glucose concentrations, etc.), bacterial culture, as well as BioFire FilmArray® Meningitis/Encephalitis (FA-ME) Panel (BioFire Diagnostics, Inc., Salt Lake City, UT) which is an automated multiplex PCR of 14 common pathogens involved in ME. Viral RNA genome was isolated from CSF, pharyngeal swabs, and stools of children positive for HPeV infection. After the synthesis of HPeV cDNA, PCR amplification of the 3' end of the VP3 region and the 5' end of the VP1 region was followed, according to the WHO surveillance guidelines (WHO 2015). CSF analysis of children with HPeV CSF infection detected no pleocytosis with a median value of 5 cells/mm3 (IQR: 1.5–10.3) and normal levels of protein (74 g/dL; IQR: 53–87.8) and glucose (40 mg/dL; IQR: 36.5–45). The median values of hospitalization days of neonates with HPeV were 7.5 days (IQR: 5.3–10.3) and of the duration of their treatment was 5 days (IQR: 2.8–8.3) (Table 1).
Genotyping of HPeV strains was carried out with Sanger sequencing. In 5/6 infants, there was available sufficient CSF quantity for further genotyping. Pharyngeal swabs and stools were also collected from these 5 neonates. Genotyping was successful in all 15 (5 CSF, 5 pharyngeal, 5 stools) different analyzed samples and HPeV-3 was the only detected genotype.
The nucleotide length of the HPeV-3 sequencing in the 5 neonates was 418 nt, 422 nt, 440 nt, 240 nt, and 239 nt. Using blast through Chromas software on the sequenced part of the VP1 region, it was revealed that 3/5 neonates were infected by a HPeV-3 strain with a complementary and inverted part of the VP1 region sequence upstream the same normal part of the VP1 region (Fig. 1A). The three strains (Genbank no. MZ277868–MZ277870) had similarities less than 95% with other recorded strains performing blast and thus, they were considered as novel strains. These three HPeV-3 strains were detected in 2018 (n = 2) and 2019 (n = 1). A sequence comparison through blast between these 3 potential recombinant strains showed 89% similarity between the first and the second, 91% between the first and the third, and 96% between the second and the third.
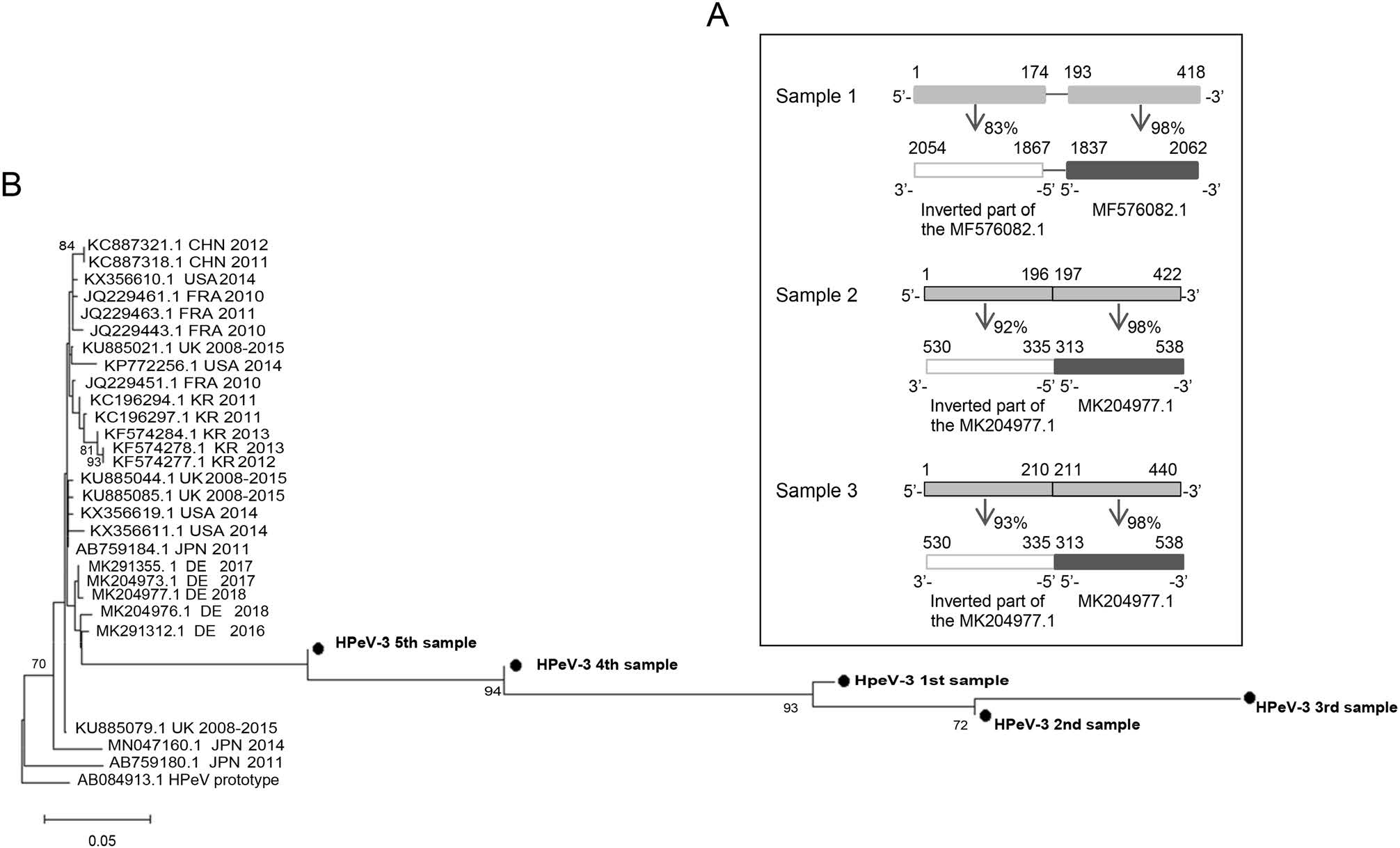
Figure 1. A Schematic representation between the three- HPeV-3 with genetic variation in the VP1 region samples of this study (light gray bars) and their most similar sequence part of the VP1 region of the recorded HPeV-3 strains (dark gray bars). The similar strains are indicated with their GenBank accession number. The white bars depict the genetic variation part of the samples of this study as an additional and inverted sequence part of the recorded HPeV-3 strains. The direction of the sequencing, the first and final nucleotide number as well as the percentage of similarity are also demonstrated. B Phylogenetic tree of HPeV-3 strains of this study with other global HPeV-3 strains based on a part of 394 nt of the VP1 region. Greek strains of this study are indicated with black circles and bold characters. The prototype AB084913 was used as an outgroup. The rest of the strains are labeled with an accession number from GenBank, the country of origin and the year of isolation. NeighborJoining method and bootstrap test of 1000 replicates were used to infer the phylogenetic trees. Only percentage values of replicate trees above 70% are shown next to branches. Notes: CHN = China, FRA = France, UK = United Kingdom, KR = Korea, JPN = Japan, DE = Germany.
Global sequences of the VP1 region of HPeV-3 isolated over the last 10 years from humans were obtained from the database GenBank (https://www.ncbi.nlm.nih.gov/genbank/). Phylogenetic tree was constructed using the Neighbor-joining statistical method and bootstrap resampling with 1, 000 replicates. The phylogenetic analysis based on partial VP1 region of HPeV-3 classified these Greek strains within the same cluster with HPeV-3 isolated in Germany in 2016, 2017, and 2018 (Fig. 1B). However, our strains were placed away from the rest, which probably indicates genetic differences in relation to the rest recorded strains in the GenBank database.
This study presented the molecular epidemiology as well as the phylogenetic and genetic characterization of HPeVs associated with pediatric CNS infections in our area. This is the first prospective study regarding HPeV CNS infection in Greek pediatric population that includes newborns and young infants. During the 3-year study period, HPeVs represented a considerable percentage of viral CNS infections, especially in neonates and young infants (11.6%). Similar results have been found in other areas, indicating that in cases of infants with ME or septiclike illness, molecular testing for HPeVs should be included in routine laboratory investigations (Vollbach et al. 2015; Chamings et al. 2019).
In the present study, the initial identification of HPeVs was made through a commercial multiplex CSF PCR platform for ME, in which HPeV detection was included and has been shown to duplicate the diagnostic yield in CNS infections compared to the control group in a previously published study from our group (Posnakoglou et al. 2020).
No pleocytosis in the CSF exam was found in infants who had HPeV CSF detection, which has also been reported in other studies (Chakrabarti et al. 2018; Habuka et al. 2020). A possible pathophysiologic explanation was presented in a recent study indicating that the immune responses in serum and CSF were distinct between HPeV-A3 and Enterovirus (EV) infected infants as it was found that higher serum levels of proinflammatory cytokine/chemokines in HPeV-A3-infected than in EV-infected patients (Habuka et al. 2020). The opposite pattern was found in CSF, with absence of CSF pleocytosis and minimal or nonexistent cytokine/chemokine response in PeV-A3-infected infants (Habuka et al. 2020). These findings might explain the more severe clinical manifestations and absence of CSF inflammation in HPeV infected infants.
The molecular genotyping was successfully performed except CSF in stool and pharyngeal samples of each HPeV positive child detecting the identical genotype. These findings could be useful for pediatricians, as CSF quantity is usually limited for PCR testing, especially in neonates and young infants (Kadambari et al. 2019).
All HPeV positive infants with CNS infection in the present study were infected by HPeV-3. HPeV-3 has been associated with aseptic meningitis, acute flaccid myelitis, and sepsis-like illness in neonates in previous reports (Vollbach et al. 2015; Zhao et al. 2016). Although in the present study all neonates with HPeV recovered without obvious acute sequelae, it has been described thatHPeV infections are competent to provoke long-term and neurodevelopmental sequelae (Berk et al. 2018; Britton et al. 2018).
The genotype HPeV-3 distinguishes from the other HPeVs because of the biannual circulation, lacking of RGD (arginine-glycine-aspartic acid) motif in the receptor binding site and the rare recombination events (Nelson et al. 2017; Sridhar et al. 2019). There are reports of novel recombinant HPeV-3 variants involved in epidemic outbreaks in infants with severe clinical manifestations (Nelson et al. 2017). In Australia, there were three consecutive epidemics with the phylogenetic analysis indicating the evolution of a recombinant HPeV-3 (Alexandersen et al. 2017; Khatami et al. 2020). The phylogenetic and genetic characterization performed for the detected HPeV-3 strains of this study unveiled the continuous evolving dynamics of this type of parechovirus.
Limitations of the present study include that it was not feasible to detect significant differences in clinical presentation or laboratory findings between the variant HPeV-3 and the other isolates due to the limited number of HPeVs cases. Moreover, only complete genome sequencing of the variant HPeV-3 genotypes could identify possible recombinations and get a deeper view of these strains.
Active enhanced surveillance of circulating HPeV genotypes with molecular methods will reveal the real clinical burden of infection, including possible outbreaks. Further phylogenetic analysis will help us identify the most common as well as emerging genotypes, owing to the frequent genetic variations or recombination events that HPeVs are incurred and contribute towards the understanding of the evolution and pathogenicity of these viruses.
HTML
-
We thank Biofire® for partial support with FilmArray® ME panels for the initial identification of HPeV.
-
Our study was conducted in accordance with the Declaration of Helsinki and the study protocol was approved by the scientific and bioethics committee of "Aghia Sophia" Children's Hospital (protocol code No: 5319). Written informed consent was obtained from the participants.


 DownLoad:
DownLoad: