











-
Since 1983, when the baculovirus expression vector was first introduced (20), the baculovirus expression vector system (BEVS) has been widely used for the expression of recombinant proteins in insect cells. BEVS has many advantages, such as the capacity for insertion of large DNA fragments, high yields of recombinant protein, and integrated post-translational modifications (17). The BEVS of two viruses, that of Autographa californica nucleopolyhedrovirus (Ac-MNPV) and Bombyx mori nucleopolyhedrovirus (BmNPV), are most widely used to express foreign proteins. More than a thousand genes have been cloned and expressed through recombinant AcMNPV and BmNPV, ranging from the components of transcription machinery to pharmaceutical products (1). However, the traditional preparation of recombinant baculovirus to express foreign genes is very time consuming, because multiple rounds of purification and amplification of recombinant viruses are needed. Recently, the newly developed Bac-to-BacTM system of AcMNPV has overcome this drawback. The AcMNPV bacmid can autonomously replicate in E. coli as a large plasmid at a low copy number, and the recombinant virus can be generated by the site specific transposition in Escherichia coli (E. coli) and used to infect insect cells. Since this system eliminates multiple rounds of purification and amplification of virus, recombinant viruses can be selected and purified within 7-10 days.
BmNPV, a member of the family Baculoviridae, is a natural pathogen of the mulberry silkworm Bombyx mori. Though the BmNPV genome is over 90% identical to the genome of AcMNPV (7), its host specificities is very narrow. Unlike AcMNPV which can infect more than 30 lepidopteran insects (2, 8), the BmNPV can only infect silkworm and its cell lines. Since it was first used to express α-interferon in 1985 (15), the BEVS of BmNPV has been used for the expression of many foreign proteins either in a cell culture system or in a insect larvae system (1).
B. mori BEVS are particularly suitable for the large-scale manufacture of foreign proteins, as the protein expression using silkworm or pupae is 10-to 100-fold higher than from B. mori cells. Silkworms are safe to the environment, and easy to breed with low cost. There is a long history of raising silkworms in China. All these make B. mori BEVS one of the most optimal systems for mass production of recombinant proteins. Several B. mori BEVS have been developed in past years (5, 10, 11, 26, 27), however, the applications were limited due to the time consuming process or low infectivity of the virus to silkworm.
In this report, we described the construction of a bacmid of BmNPV, BmBacJS13, using in vivo recombination. To study the infectivity of the bacmid, thepolyhedrin gene was inserted into the bacmid generating recombinant virus BmBacJS13-ph. The infectivity of BmBacJS13-ph was demonstrated by growth curve analysis of budded viruses and bioassay in B. mori larvae. The results indicated that BmBac-JS13-ph is a functional virus with similar infectivity to that of wt BmNPV. The results also showed that infective recombinant viruses could be generated with BmBacJS13 by site specific transposition in E. coli, indicating a functional Bac-to-Bac system of BmNPV was constructed.
HTML
-
Larvae of Bombyx mori, were reared on an artificial diet at 27℃ (4) and third or fifth instars larvae were used in the experiments. BmN cells were cultured in TC-100 (JRH) insect medium supplemented with 10% fetal bovine serum (GIBCO/BRL) at 28 ℃ using standard techniques (17). The wt BmNPV Shaanxi strain, collected in Shaanxi province, China, were propagated on silkworm larvae (3).
-
According to the sequence of the BmNPV T3 strain (GenBank accession number L33180), a 1.5 kb segment upstream of polyhedrin gene was amplified by PCR from the wt BmNPV genome DNA using primers(5'-GGGCCGCGGGCGTAGAGATTCGACGAAAGC-3' and 5'-GCGCTGCAGATAATTACAAATAGGATTGAGGCC-3') and cloned into the Sac Ⅱ and Pst Ⅰ sites of pBluescriptKS Ⅱ+ (Stratagene). A 1.7 kb segment downstream of the polyhedrin gene was PCR amplified using forward primer (5'-GGGGAATTCCCTGAGGTAAGCGTTAGATTCTGTGCGTTTG-3') with EcoR Ⅰ and Bsu 36Ⅰ sites, and reverse primer (5'-GCGGGTACCTGACGAATCGTAAATATGAATTCTGT -3') with Kpn Ⅰ site. The product was cloned into the plasmid containing the upstream segment to generate pKS-USR-DSR. The mini-F replicon, kanamycin resistance gene and the Tn7 target sites were cut out as an 8.6 kb Bsu36Ⅰ fragment from plasmid pBAC-Bsu36Ⅰ (18) and inserted into the Bsu36Ⅰ site of pKS-USR-DSR to generate transfer vector pBmTV (Fig. 1).
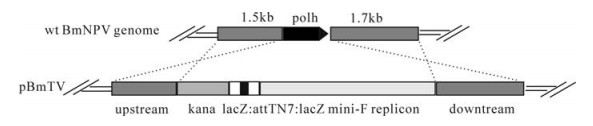
Figure 1. Schematic of transfer vector pBmTV. The 1.5kb upstream sequence and 1.7kb downstream sequence are homologous arms, through recombination the polh was replaced with kanamycin cassette, lacZ: attTN7: lacZ and mini-F replicon.
-
Newly molted fifth instar larvae were co-trans-fected with 20 μL DNA mixture per larva (including 0.45μg linearized plasmid pBmTV DNA, 0.15 μg wt BmNPV viral DNA and 3μL lipofectin (Invitrogen)) through subcutaneous injection (25). The haemolymph of the larvae were collected at 4 days post transfection and budded viruses (BVs) were retrieved from the supernatant of haemolymph after centrifuging. The DNA of BVs was extracted and transformed into E. coli DH10B cells (GIBCO/BRL), and colonies were selected in the presence of kanamycin and X-Gal. The colonies were further analyzed by PCR and REN digestion. One of the positive clones, BmBacJS13 which had similar REN profiles to that wt BmNPV, was chosen for further analysis.
-
The Polyhedrin gene of BmNPV was amplified by forward primer (5'-GCGGGATCCTGTCGACAAGCTCTGTCCGTTT-3') with BamHI (underlined), and reverse primer (5'-GGGGAATTCTTAATACGCCGGACCAGTG-3') with EcoR Ⅰ (underlined). The BamH Ⅰ site of the fragment was blunted and the fragment was inserted into Bst1107 Ⅰ and EcoR Ⅰ sites of pFast bac Dual (Invitrogen) to generate pFast-DUAL-ph. The pFast bac Dual-ph was then used to produce a recombinant BmBacJS13-ph via transposition in E. coli according to the Bac-to-BacTM system manual. The bacmid DNAs of the recombinant were extracted and were used to transfect the BmN cells.
-
Bacmid DNA of BmBacJS13-ph was isolated by methods developed for large plasmids (instruction manual of Bac-to-BacTM system/Life Technologies). The DNA was used to transfect BmN cells with lipofectin, and genomic DNA of wt BmNPV was used as a control. The supernatant was collected from the transfected BmN cells at 96 hrs post transfection (h.p.i.) and was used to infect BmN cells. The supernatants of the infected cells were collected at 96 h.p.i and TCID50 was determined using end-point dilution method.
-
BmN cells (1×106) were infected with wt BmNPV or BmBacJS13-ph with a multiplicity of infection (MOI) of 5. The infected cells were harvested at 72 h.p.i. The samples were processed for electron microscopy examination as described by Van Lent et al. (22).
-
BmN cells were infected with BV of BmBacJS13-ph or wt BmNPV at an MOI of 5. At the appropriate time points post infection, the supernatants were collected and the titers were detected using the end-point dilution method, Each virus infection was repeated five times and the data were statistically analyzed with one-way ANOVA (SPSS); polyhedral inclusion body (PIB) was used as the marker during the assay.
-
The third instars of B. mori larvae were used in oral infection assays to determine the infectivity of BmBacJS13-ph and wt BmNPV. The larvae (n=50 per viral dose) were fed with artificial diet which containned different concentrations of viruses (3×107 PIB/mL, 107 PIB/mL, 3×106 PIB/mL, 106 PIB/mL, 3×105 PIB/mL and 105 PIB/mL, respectively) for twenty-four hrs. Then the larvae were transferred to fresh diet in plates and reared at 27 ℃ to investigate the mortality. The infectivity of BmBacJS13-ph and wt BmNPV was determined by probit analysis with SPSS10.0 and LC50 values of the viruses were further compared with a two side's z-test (21).
Insect, virus and cell line
Construction of transfer vector
Recombination and identification of the bacmid in vivo
Construction of BmBacJS13-ph
Transfection of BmN cells
Electron microscope
Comparison of the growth curves between wt BmNPV and BmBacJS 13-ph
Bioassay
-
After the construction of transfer vector pBmTV (Fig. 1), the pBmTV DNA was co-transfected with BmNPV DNA into newly molted fifth instar larvae. The hemolymphs of the larvae were collected 4 days post transfection, and BV DNAs were retrieved to transform E. coli DH10B cells. Colonies were chosen randomly and bacmid DNAs were digested with Hind Ⅲ. By comparison with the wt BmNPV profiles, several variations were found in different colonies (data not shown). One of the colonies, BmBacJS13, which had similar REN profiles to that of wt BmNPV, was analyzed with additional restriction enzymes. As show n in Fig. 2, co mparison with the w t BmNPVindicates that the Xba Ⅰ profile of BmBacJS13 lacked B (19.3 kb), D (15.8 kb), G (7.6 kb) and H (6.3 kb) bands, and contained five additional bands, D+H (22.2 kb), B' (19.0kb), G' (9.1 kb), 4.9 kb and 1.6 kb. Due to the deletion of the polyhedron gene, the B band in wt BmNPV became a shorter band B' in BmBacJS13. The 4.9 kb and the 1.6 kb bands were from the inserted 8.6 kb Bsu36Ⅰ fragment which contains three Xba Ⅰ sites. The remaining Bsu36Ⅰ fragment (2.1kb) was linked with the remainder of G the fragment (7.0 kb) to generate G'(9.1 kb). The D+H band in BmBacJS13, h o wever, was a sub-molecular band of the D and H bands in the wt BmNPV. These were the expected changes for the bacmid. Therefore BmBacJS13 was constructed correctly and was used for further investigation
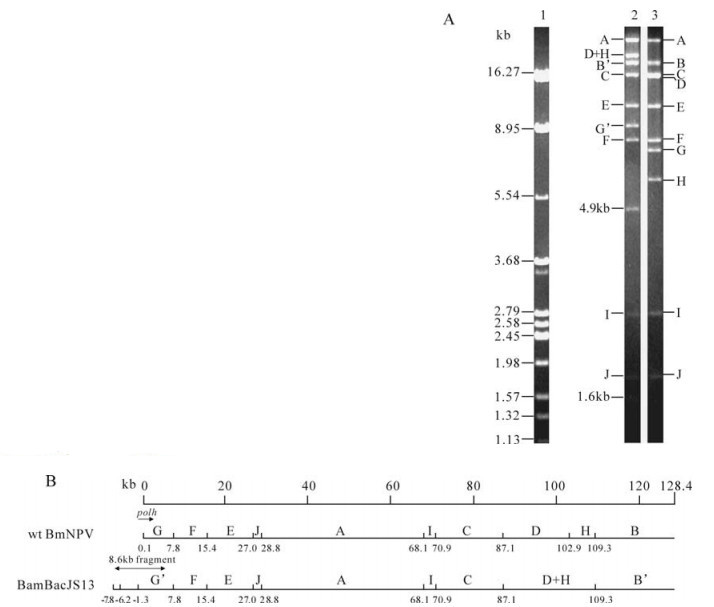
Figure 2. Xba Ⅰ digestion profiles and linearized physical maps of BmBacJS13 and wt BmNPV. A. Xba Ⅰ digestion profile of BmBacJS13 and wt BmNPV. 1, λ DNA digested by BamH Ⅰ, EcoR Ⅰ and Hind Ⅲ; 2, BmBacJS13; 3, wt BmNPV. B. Linearized Xba Ⅰ physical maps of BmBacJS13 and wt BmNPV. The restriction sites are indicated in kb from the zero point. The genome size in kb is shown on a scale at the top. The location of polh and 8.6kb Bsu36Ⅰ fragment are shown.
-
To test the oral infectivity of BmBacJS13, the polyhedrin gene was repaired into the bacmid, generating BmBacJS13-ph. The BmBacJS13-ph was identified to be correct by PCR and REN analysis (data not shown), and transfected into BmN cells to generate BVs.
BmN cells were infected with BmBacJS13-ph at an MOI of 5, occlusion bodies were observed in the nucleus of cells by optical microscopy from 48 hrs p.i, and the detachment of the cells were observed at the same time. These cytopathic effect (CPE) were similar to that of wt BmNPV infected cells. Electron microscopy analysis showed that the occlusion bodies were formed in the nuclei. The polyhedra and ODVs of BmBacJS13-ph had a similar shape and size as those of wild type BmNPV (Fig. 3).
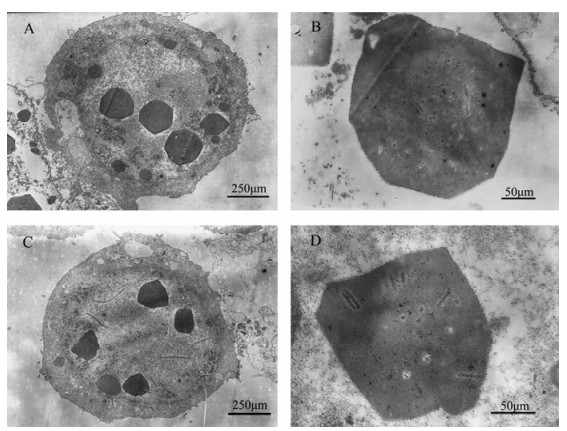
Figure 3. EM pictures of BmN cells infected with wt BmNPV (A, B) and BmBacJS13-ph (C, D).
One-step growth curve analysis indicated that BmBacJS13-ph BV had similar replication dynamics to that of wt BmNPV (Fig. 4). The titers of the two viruses were very similar during the early infection, but the titers of BmBacJS13-ph were slightly lower than that of wt BmNPV during 24 to 96 h.p.i. Statistical analysis indicated that the differences were not significant except those for 72 h.p.i..
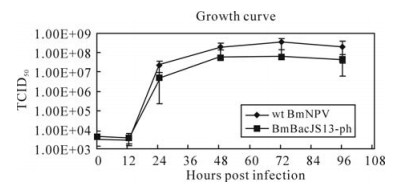
Figure 4. Growth curves for BmBacJS13-ph and wt BmNPV. BmN cells were infected with BmBacJS13-ph or wt BmNPV at an MOI of 5. The supernatants were harvested at different times post infection and the BV titers were analyzed by EPDA. The average titer from five independent TCID50 assays were shown with the bars indicating standard errors.
The bioassay result showed that LC50 of BmBac-JS13-ph against 3th larvae was 3.9x106 PIB/ml, which was not significantly different from of that of wt Bm-NPV (3.7x106 PIB/ml) (z=0.3276, P > 0.05) (Table 1).

Table 1. LC50 of wt BmNPV and BmBacJS13-ph in early third instar B. mori larvae
Identification of the bacmid BmBacJS13
Biological activity of BmBacJS13-ph
-
A functional Bac-to-Bac system should possess the following characteristics: first, the bacmid should be able to autonomously replicate in the bacteria; second, a foreign gene can be inserted into the bacmid via transposition, and lastly, the backbone virus must have similar biological properties as the wt virus, in both cells and insects. Our study showed that BmBacJS13 could autonomously replicate in the bacteria, and a foreign gene (ph) was introduced into the Tn7 site of BmBacJS13 with the aid of the helper plasmid. The recombinant virus BmBacJS13-ph had similar replication dynamics to that of wt BmNPV and bioassay results showed that its in vivo infectivity was similar to that of wt BmNPV. Therefore a functional Bac-to-Bac system of BmNPV has been constructed, and BmBacJS13 can be used to construct BmNPV recombinants for expressing foreign proteins. During our experiments, a similar Bac-to-Bac system of BmNPV was reported by Motohashia et al (16). In their system, a bacmid of the BmNPV T3 strain was constructed, and it could express enhanced green fluorescence gene (egfp) in larvae and pupae. As we have used a local strain from China (Shaanxi strain), it would be interesting to compare the properties between BmBacJS13 and the T3 bacmid.
The Bac-to-Bac system can also be used for functional genomics studies of BmNPV. Bacmids have been widely used to delete genes by site-specific recombination in E. coli in AcMNPV and Helicoverpaarmigera NPV (6, 9, 13, 12, 14, 19, 24, 23).. The target genes were deleted and subsequently repaired by transposition to determine the function of the genes in viral life cycle. Currently we are studying the functions of several BmNPV genes using BmBacJS13 and the results will be reported in the future.


 DownLoad:
DownLoad: