












-
Dear Editor,
The coronavirus disease 2019 (COVID-19) pandemic presents a severe threat to public health and the global economy and the direct origin of the causative agent severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) remains unresolved (WHO 2020). The outbreak in China was successfully contained by the implementation of a strict quarantine strategy, however sporadic outbreaks still occurred and were associated with trade markets in Beijing and Dalian cities. The COVID-19 outbreak in Beijing was traced to contaminated salmon meat sold at the Xinfadi market, Beijing, China (ChineseCDC 2020), causing by a viral strain belonging to the B1.1 lineage, which firstly identified and mainly circulated in Europe, indicating that the strain had transferred from another country (Yang et al. 2020). Later, the Chinese Center for Disease Control and Prevention detected the nucleic acid of SARS-CoV-2 from over ten samples of imported frozen shrimp and chicken meat products in several cities in China (Chen and Duan 2020). Additionally, outbreaks associated with slaughterhouses and meat-packing factories in Portugal, Ghana, UK, and Australia were reported (Waltenburg et al. 2020). The spread of SARS-CoV-2 from workers to food products is conceivable. Hence, food safety and the analysis of the potential relation between food and the outbreaks of COVID-19 are of great significance. Here, we aimed to determine the stability of SARS-CoV-2 on meat surfaces in the setting that simulates the cold chain transportation, in order to provide scientific advice for transmission control.
Pork, beef, and frozen salmon meat were bought from supermarkets (Wuhan & Kunming) and cut into cubes that weighed approximately 1-2 g per cube. The stock SARS-CoV-2 (IVCAS 6.7512 in Wuhan and NMDCN000HUI in Kunming) (Zhou et al. 2020) was diluted with Dulbecco's modified Eagle's medium (DMEM) (Gibco) to 103 or 104 TCID50/mL. The meat was immersed in the DMEM containing SARS-CoV-2 at 103 or 104 TCID50/mL for 10 s. Later, these cubes were transferred into a 6-well plate and then stored at 4 ℃ or - 20 ℃ to simulate the temperature of cold chain transportation. Meat samples were taken out at different time points, and each cube was transferred to 24 wells plate and rinsed 20 times with 1 mL of DMEM. Each type of meat sample was tested in two to six technical replicates at each time point (Table 1). Then, 140 µL of washing medium was used to extract viral RNA using the Viral RNA kit (Roche, Germany), and the viral RNA extracted in the washing medium was amplified using quantitative reverse-transcription polymerase chain reaction (qRT-PCR) targeting the spike genes of SARS-CoV-2 (Mabsky Biotech Co., Ltd., China). The limit of detection (LOD) of the kit was 610 viral RNA copies/mL. Additionally, 200 µL of the washing medium was used to infect Vero E6 cells (ATCC no. CRL-1586) in a 24-well plate. Three days later, the supernatant in each well was collected to detect the viral RNA by qRT-PCR; meanwhile, the cells were washed thrice with 1× phosphate buffered saline (pH 7.4) and then fixed with 4% paraformaldehyde to detect the viral nucleocapsid protein using the indirect immunofluorescence assay (IFA) as described previously (Zhou et al. 2020). The viral isolation was considered successful from either qRT-PCR or IFA-positive test results of the infected cell or the culture supernatant. Meat samples were collected at 0, 12, 24, 36, 48, 72, 144, and 216 h after stored at 4 ℃, and at 0, 1, 2, 3, 4, 5, 6, 7, 9, 15, and 20 days after stored at -20 ℃ (Table 1).
103 TCID50/mL 104 TCID50/mL Pork Beef Salmon Pork Beef Salmon 4 ℃ 0 h 5/6 5/6 3/6 6/6 6/6 6/6 12 h 4/6 3/6 6/6 6/6 6/6 6/6 24 h 4/6 4/6 4/6 6/6 6/6 6/6 36 h 4/6 3/6 3/6 6/6 6/6 4/6 48 h 3/6 5/6 2/6 6/6 6/6 5/6 72 h 6/6 6/6 3/6 6/6 6/6 5/6 144 h / / / 2/2 2/2 2/2 216 h / / / 2/2 1/2 1/2 - 20 ℃ 0 D 5/6 5/6 4/6 6/6 5/6 6/6 1 D 4/6 6/6 3/6 6/6 6/6 6/6 2 D 5/6 6/6 5/6 6/6 6/6 6/6 3 D 5/6 5/6 3/6 5/6 6/6 6/6 4 D 6/6 6/6 2/6 6/6 6/6 5/6 5 D 6/6 6/6 2/6 6/6 6/6 6/6 6 D 0/3 2/3 1/3 3/3 3/3 3/3 7 D 2/3 3/3 2/3 2/3 3/3 3/3 9 D 1/2 0/2 1/2 1/2 2/2 0/2 15 D 1/2 1/2 2/2 2/2 1/2 1/2 20 D 0/2 0/2 1/2 1/2 1/2 1/2 Virus isolation successful repeats/total repeats. Table 1. Isolation rates of SARS-CoV-2 from washing liquids of different meat samples.
From the samples containing low viral titers stored at 4 ℃, viral RNA could be detected in the washing fluids of all the samples until 72 h post-infection (hpi) (Fig. 1A). The average number of viral RNA copies was higher in pork meat samples than in beef and salmon meat samples. The number of viral RNA copies in low titers-treated salmon samples was lower than the LOD number of the kit. In samples containing high viral titers stored at 4 ℃, viral RNA could be detected in the washing fluids of all the samples until 216 hpi, and the viral RNA copies in three types of meat samples show no significant difference (Fig. 1B). The average number of viral RNA copies showed no decline over time, both in the low and high viral titer groups. Regarding viral isolation, live viruses could be isolated on Vero E6 cells from samples stored at 4 ℃ at all the time points, but not from all of the technical replicates (Table 1). The viral isolation rates in three types of meat samples show no significant difference. At high viral concentration, the virus was successfully isolated from all washing liquid samples of pork and beef until 6 days post-infection (dpi). However, the isolation rate was lower from the samples of salmon meat. At 9 dpi, the highest live-virus isolation rate was measured from the samples of pork meat.
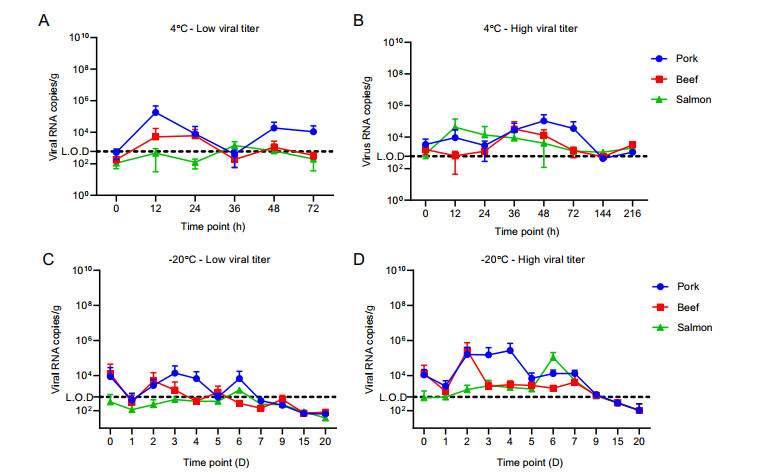
Figure 1. Number of viral RNA copies in the washing liquid of different meat samples. The washing medium was used to extract viral RNA using the Viral RNA kit (Roche, Germany), and the viral RNA extracted in the washing medium was amplified using quantitative reverse-transcription polymerase chain reaction (qRT-PCR) targeting the spike genes of SARS-CoV-2 (Mabsky Biotech Co., Ltd., China). The LOD of the kit was 610 copies of RNA/mL. LOD, limit of detection. A Viral RNA copies in the washing fluids of low viral titers stored at 4 ℃; B Viral RNA copies in the washing fluids of high viral titers stored at 4 ℃; C Viral RNA copies in the washing fluids of low viral titers stored at - 20 ℃; D Viral RNA copies in the washing fluids of high viral titers stored at - 20 ℃.
In meat samples containing low viral titers stored at - 20 ℃, viral RNA could be detected in the washing fluids of all the samples until 20 dpi at the experiment ending point (Fig. 1C). The average number of viral RNA copies was lower in the salmon meat sample than in the beef and pork meat samples. The number of viral RNA copies in all three meat samples was lower than the LOD number of the kit after 7 dpi. In meat samples containing high viral titers stored at - 20 ℃, viral RNA could be detected in the washing fluids of all samples until 20 dpi (Fig. 1D). The viral RNA copies in three types of meat samples show no significant difference. The number of viral RNA copies was lower than the LOD number of the kit after 9 dpi. Regarding viral isolation, live viruses could be isolated on Vero E6 cells from all of the samples, but were not successful in all the technical replicates (Table 1). The virus was successfully isolated from the washing fluid samples of all the pork and beef meat samples until 6 dpi; however, the isolation rate decreased over time.
In this study, we tested three types of meat stored under two different storage temperatures after inoculation with two different viral concentrations. Viral RNA and live viruses could be detected or isolated from the surface of meat samples collected at different time points. Viral RNA copies were lower on salmon compared with pork and beef, which may be caused by the different texture and fat content of the meat. The viral isolation rate showed no difference under 4 ℃ and - 20 ℃ before 3 dpi; however, the viral isolation rate of the frozen meat samples (- 20 ℃) was higher than that of the fresh storage meat samples (4 ℃) after 3 dpi. Recently, a study published in bioRxiv tested the viral stability in chicken, pork, and salmon meat samples, and found that the viral titer does not reduce at 4 ℃, - 20 ℃, and - 80 ℃, respectively, for 3 weeks (Fisher et al. 2020). However, the viral concentration used in the study was as high as 3 × 106 TCID50/mL, which is significantly higher than the viral titers present in clinical respiratory samples (1 × 103 to 1 × 104 TCID50/mL) that correspond to 107-108 viral RNA copies/mL (Pan et al. 2020; To et al. 2020; Zhang et al. 2020). In our study, we used two viral concentrations and tried to simulate the viral titers present in patient respiratory samples. For a better understanding of the survive time of live virus on the surface of different types of meats, we will carry out a study for a long-stored period in the future.
Generally, compared to those of the lower viral concentration group, samples containing a higher viral concentration had a higher number of viral RNA copies and higher viral isolation rates. However, the viral RNA detection rate is not consistent with the rate of viral isolation as the two methods have different sensitivity. Other groups tested the stability of SARS-CoV-2 at different temperatures and on different surfaces and found that SARS-CoV-2 is sensitive to high temperatures and dry conditions, but it is more stable at low temperatures (Chin et al. 2020; Fisher et al. 2020; van Doremalen et al. 2020). Given the stability of SARS-CoV-2 at low temperatures, there is a risk of SARS-CoV-2 transmission from food products that are a part of the cold chain. Therefore, it is important to quarantine the staff working in food production, packaging, and transportation. We also suggest making necessary hygiene treatment and disinfection processes for food products, before consuming products transported by the cold chain.
HTML
-
This work was supported by the Key Program of Chinese Academy of Sciences (KJZD-SW-L11 to Z.L.S.), the Strategic Priority Research Program of the Chinese Academy of Sciences (CAS) (XDB29010102 and XDA19090118 to Y.H.B.), COVID-19 emergency project of CAS (2020YJFK-Z-0149), the National Key R & D Program of China (2020YFC0842000 to Y. T. Z.), the National Natural Science Foundation of China (NSFC32041010 to Y.H.B.), the NSFC Outstanding Young Scholars (31822055 to Y.H.B.), and Youth Innovation Promotion Association of CAS (2017122 to Y.H.B., 2019328 to X.L.Y.).
-
The authors declare that they have no conflict of interest.
-
This article does not contain any studies with human or animal subjects performed by any of the authors.

 DownLoad:
DownLoad: