











-
Hepatitis B virus (HBV) is a serious public health problem. The best antiviral therapy at present is IFN and nucleoside analogues. But high resistance mutations to nucleoside analogues and low response to IFN makes it difficult to eradicate hepatitis B. Given the widespread non-response to exogenous IFN-α, novel therapeutic approaches that either augment or complement the antiviral activity of IFN-α would be very beneficial.
Several factors could be associated with the low response to IFN, such as age, sex, HBV DNA levels and transaminase (ALT) level in hepatitis B patients. Recently, some studies have shown that when compared with responder's liver tissue, non-res-ponders have increased expression of a number of IFN-sensitive genes (3). The expression of a subset of 8 genes accurately predicted treatment response in more than 90% of patients as well as viral load, ALT level, or hepatic fibrosis. Two of the 8 genes, UBP43 and IFN-sensitive gene ISG15, are linked biochemi-cally. ISG15 is a ubiquitin-like molecule that is post-translationally attached to the lysine residues of more than 150 target proteins (6, 17). Protein targets include other IFN-stimulated genes, in addition to proteins in a diverse set of unrelated pathways. However, the function of the ISG15 conjugation remains unknown. UBP43 is a ubiquitin-specific protease that cleaves the ubiquitin-like (and IFN-induced) ISG15 protein from its cellular targets in vitro(12). Together these data suggest that non-responder patients have a disordered host IFN response, and the up-regulation of UBP43 can be correlated with non-response to IFN treatment.
Multiple lines of evidence previously have linked ISG15, UBP43, and the cellular response to IFN. Both genes are induced by type Ⅰ IFN in many cell types (4, 5, 13), and cells isolated from UBP43 knockout mice are hypersensitive to IFN, with prolonged Jak–Stat signaling (14).These proteins also have been linked to the innate immune response to viruses. Overex-pression of ISG15 enhances the antiviral activity of IFN against human immunodeficiency virus and Sindbis virus replication in vitro(10, 11, 13). In-fluenza B virus NS1 protein binds ISG15 and prevents host protein ISGylation, a function that correlates with influenza B resistance to IFN (15).Based on these data, we hypothesized that UBP43 may be a critical determi-nant of the human IFN antiviral response to HBV.
HTML
-
Restriction enzymets Pst Ⅰ, EcoR I, BglⅡ, Sal Ⅰ, T4 polynucleotide kinase and T4 DNA ligase were obtained from Takara (Japan). Plasmid purification kits were purchased from Omega (USA). Lipofec-tamineTM was obtained from Invitrogen and the ELISA kit from the Dakewei Company (China). DMEM and fetal calf serum were purchased from Hyclone and Sijiqing Biotech Company (China) respectively. pSUPER vector was kindly provided by Professor Xin-hua Feng (Department of Molecular and Cellular Biology, Baylor College of Medicine Houston, United States). HepG2.2.15 cells and Escherichia coli DH5α were preserved in our laboratory.
-
shRNAs were cloned downstream of the human H1 promoter in the vector pSUPER as described previously (1). The target sites for siRNA were recommended by the website (www.ambion.com and www.dharmacon.com). The selected sequences were submitted to a BLAST search to avoid targeting to the Human genome. The sequence was as follows: 5'-GATCTCCGGAATTCACAGACGAGAAATTTTCAAGAGAAATTTCTCGTCTGTGAATTCCTTTTTG-3', 5'-TCGACAAAAAGGAATTCACAGACGAG AAATTTCTCTTGAAAATTTCTCGTCTGTGAATTCCGGA-3' and irrelevant sequences was 5'-GATC TCCATGGCTTAAGCAAAGTGAAACTTCAAGAGAGTTTCACTTTGCTTAAGCCATTTTTTG-3', 5'-TCGACAAAAAATGGCTTAAGCAAAGTGAAACTCTCTTGAAGTTTCACTTTGCTTAAGCCATGGA-3'. The phosphorylation, regeneration and annealing of single siDNA were performed as follows: after mixture of water, polynucleotide kinase buffer, single siDNA, ATP and T4 polynucleotide kinase, then 37 ℃ for 45 min, 95 ℃ for 5 min, and 55℃ for 10 min. The product was ligated into purified pSUPER plasmid from 1% agarose gel which was enzymatic digested with Sal Ⅰ and Bgl Ⅱ. The recombinant plasmid was used to transform Escherichia coli DH5α cells. The positive clones were selected and verified by DNA sequence.
-
HepG2.2.15 cells, a human hepatoblastoma HepG2 cell line stably transfected by the HBV genome, were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum. Then HepG2.2.15 cells were transfected in 6-well plates using LipofectamineTM. The cells were divided into 3 groups, namely the interference group (transfected with psiUBP43), the negative control group (trans-fected with pSuper plasmid by inserting a nonspecific sequence, Nonspecific sequence means that the nucleotide bases were the same as the target seqeuence but the order was different.So we also refer to the negative control as the irrelevant control.) and the blank control group (without transfection). Transfection of cells was performed with lipofectamine following the manu-facturer's guidelines. The plate was incubated at 37 ℃ with 5%CO2 for 4 h, after which 2mL DMEM medium containing 10% FBS were added into the well, with a final concentration of IFN equal to 3000IU/mL. After further incubation for 48 h, the cells were taken for observation.
Total RNAs were extracted 48h after transfection. The cells were collected from wells in each group for total RNA extraction with Trizol according to the manufacturer's instructions. After being dissolved in DEPC, the RNA was accurately quantified with an ultraviolet (UV) spectrophotometer.
-
The primers for GAPDH, UBP43 were as follows: UBP43-s: 5'-CAGACCCTGACAATCCACCT-3'; UBP-43-as: 5'-AGCTCATACTGCCCTCCAGA-3'; GAPDH -s: 5'-CGGATTTGGTCGTATTGGG-3'; GAPDH-as: 5'-CTCGCTCCTGGAAGATGG-3'. We used a RQ-PCR assay based on Sybergreen fluorescence methodology to quantify the full range of UBP43 mRNA copy numbers (2, 9). The number of PCR cycles required to reach the fluorescence threshold was the cycle threshold (Ct). The Ct value for each sample was proportional to the log of the initial amount of input cDNA. By plotting the Ct value of an unknown sample on the standard curve, the amount of target sequence in the sample could be calculated. To normalize the UBP43 mRNA expression for sample to sample differences in RNA input, RNA quality, and reverse transcriptase efficiency, we amplified the housekeeping gene GAPDH. From their respective standard curves, we obtained the copy numbers of GAPDH and UBP43. The ratio between copy numbers of UBP43 and GAPDH represented the normalized (nUBP43) for each sample and could be compared with that of other samples and were calcu-lated according to nUBP43 = (UBP43 mRNA copies sample/GAPDH mRNA copies sample) ×10 000 (1).
-
Following the protocol provided with the antigen detection kit, the concentration of HBsAg and HBeAg in the supernatant were detected by ELISA. The results were presented as S/CO values. Extraction of HBV DNA replicative intermediates and Southern blotting were performed as previously described (8, 17).
Experiment materials
pSuper plasmid construction
Cells culture, preparation of total RNAs
Assay of UBP43 mRNA expression after RNA interference
ELISA and Southern blot
-
The result of 2.5% agarose gel electrophoresis showed that the plasmid psiUBP43 and irrelevant control cut by EcoR I and Sal Ⅰ was about 60 bp longer than the pSUPER blank control as shown in Fig. 1 and was verified by sequencing. Because the psimIFNAR1 and irrelevant plasmid had inserted shRNA into plasmid pSUPER, so they were about 60 bp longer.
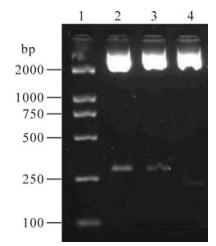
Figure 1. Agrose gel electrophoresis of double enzyme digestion of plasmid. 1, DL2000 marker; 2, psiUBP43 digested by EcoR I and Sal Ⅰ; 3, pSuper plasmid by inserting a nonspecific sequence; 4, pSuper digested by EcoR I and Sal Ⅰ.
-
We found that the expression of UBP43 was decreased compared to the control. For each experi-mental sample, the amount of UBP43 and GAPDH was determined from the appropriate standard curve. Then, the UBP43 amount was divided by the GAPDH amount to obtain the nUBP43 ratio. The nUBP43 ratio was significantly lower (P < 0.05, also significant after Bonferroni correction for multiple comparisons) in the interference group than in the negative control and blank control (Fig. 2).
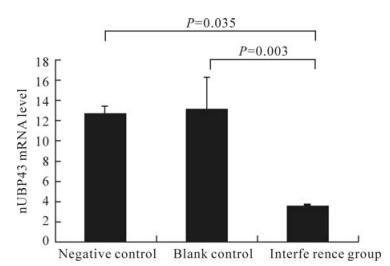
Figure 2. Realtime fluorescence quantiative RT-PCR assay of UBP43 mRNA (n=3 Mean±SD). Negative control (with irrelevant pSuper) versus interference group (transfected with psiUBP43) (P=0.035) and blank control (without plasmid) versus interference group (P=0.003). nUBP43 was calculated according to equation 1.
-
The HepG2.2.15 cells were divided into three groups: the interference group, the negative control and the blank control. They were treated with IFN (3000IU/mL) for 48h; the HBeAg and the HbsAg were detected in the medium by ELISA and HBV DNA isolated from treated HepG2.2.15 cells was measured by Southern Blot. The secretion of HBeAg, but not HBsAg, was significantly suppressed compared to controls (Fig. 3). and it was found that the HBV replication was significantly inhibited compared to the controls (Fig. 4).
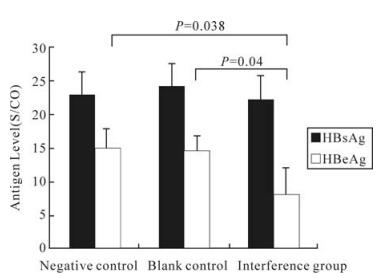
Figure 3. Inhibition of viral gene expression by shRNA in sequence specific fashion. After transfection of HepG2.2.15 cells with shRNA expression plasmids and addition of IFN, the HBeAg expression was substantially suppressed, but not for HBsAg and HepG2.2.15 cells which were more sensitive to IFN by psiUBP43 relative to blank control (without plasmid) and negative control (with irrelevant pSuper) (P < 0.05).
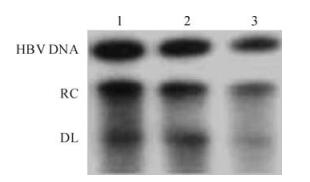
Figure 4. Inhibition of viral replication by IFN after trasfection of shRNA. After transfection of HepG2.2.15 cells with psiUBP43, the intracellular viral replicative intermediates were detected by Southern blotting. RC DNA: relaxed-circular DNA; DL: Double-stranded linear DNA. 1, Negative control (with irre-levant pSuper plasmid); 2, Blank control (without plasmid); 3, Inter-ference group (transfected with psiUBP43).
pSuper plasmid construction
Effect of shRNA on UBP43 mRNA expression
Effects of silencing UBP43 on secreted HBsAg and HBeAg
-
In a recent study, it was found that expression of UBP43 was disordered in the pretreatment liver tissue of patients with chronic HCV who do not respond to subsequent treatment with pegylated IFN-2-α and ribavirin (3). The study also showed that silencing the UBP43 increases the antiviral activity of IFN against hepatitis C virus infection. So, we hypothesized that silencing UBP43 might enhance the antiviral activity of IFN-α to inhibit HBV replication.
The pSUPER plasmid used in this study is a good tool for generating small hairpin RNAs (shRNAs) to interfere with the expression of the targeted gene. First, we constructed the recombinant shRNA plasmid (psiUBP43) by inserting the oligonucleotides down-stream of the H1 promoter. When we transfected psiUBP43 into the HepG2.2.15 cells, it significantly suppressed the expression of UBP43 compared with negative or blank controls. We then tested the effect of UBP43 on IFN-α biological activity. We show that UBP43 also plays an important role in the anti-HBV type Ⅰ IFN response using HepG2.2.15 cells. The silencing of UBP43 altered the cellular response to IFN-α, resulting in increased inhibition of gene expression (HBeAg) and HBV replication. The increased biochemical effect of IFN-α was mirrored by a potent inhibition of HBV DNA in vitro. So our results suggest that UBP43 plays an important role in HBV pathophysiology.
UBP43 may have a broader role in human disease. In animal models, UBP43 knockout mice are resistant to otherwise fatal intracerebral infection by lym-phocytic choriomeningitis virus and vesicular sto-matitis virus. This effect was associated with decreased viral replication and increased cellular ISGylation by ISG15 (16).However, the mechanism by which UBP43 modulates the antiviral activity of IFN remains unclear. Most scientists agree that it play an important role in the negative regulation of IFN signaling. Because UBP43 is itself induced by IFN-α, it appears to be a critical regulator in a classic negative feedback loop. UBP43 would appear to regulate a process upstream of STAT1 phosphorylation, such as IFN-α– receptor turnover or the interactions between the receptor and signaling molecules. Prolonged STAT1 activation is associated with a general increase in IFN-stimulated gene expression in UBP43-silenced cells.
We believe that the question of how UBP43 contributes to IFN-α antiviral activity in human HBV is a critical one. Not only is IFN-α the major component of antiviral therapy for chronic HBV infection, but non-response to IFN-α is an important and unresolved issue in HBV treatment. IFN-α has been shown to target multiple stages of viral life cycles, notably translation, but also viral entry and capsid assembly (7). The HBV infectious cell culture system used in this study models the entire viral life cycle, and therefore provides the most advanced system for studying the antiviral activity of IFN-α against HBV. UBP43 silencing in this system produced a robust and consistent enhancement of the ability of IFN-α to inhibit HBV replication.
There fore, enhancing the antiviral activity of IFN by modulation of UBP43 may represent a new strategy for improving responses to HBV treatment.


 DownLoad:
DownLoad: