














HTML
-
A key characteristic of herpes simplex viruses 1 and 2 (HSV-1 and HSV-2) is that once virus entry into the cells, viral DNA is transported to neurons of sensory ganglia in which they either express immediate early viral regulatory proteins to initiate viral lytic replication or express an 8.3–9 kb RNA designated as the latency associated transcript precursor (LAT-P) to establish a latent infection (Nahmias and Roizman 1973; Roizman and Sears 1987). The LAT-P accumulates at low levels in latently infected neurons, whereas the 2- and 1.5-kb LAT introns processed from LAT-P are abundant and stable accumulated in nuclei (Wagner et al. 1988; Farrell et al. 1991). LAT is the major viral transcript detected in neuronal nuclei by in situ hybridization (Stevens et al. 1987). One of the many questions posed by HSV latency is the role of LAT in silencing the transcription of viral genes essential for the initiation of viral replication (Nicoll et al. 2016).
The first step in the replication of HSV-1 is the transcription of a set of viral genes known as a or immediate early genes. The a proteins perform essential regulatory functions which are necessary for viral DNA synthesis and capsid assembly (Honess and Roizman 1975). An important member of this group is ICP0 (Cai and Schaffer 1992). One potential mechanism by which LAT-P could interfere with the expression of viral genes is by sequestering ICP0 mRNA and thereby blocking accumulation of ICP0 (Garber et al. 1997; Mador et al. 1998). In brief, ICP0 mRNA is in its entirely complementary to the LAT-P. LAT is derived from the 5' domain of LAT-P and complementary to less than 1 kb of the 3' terminal sequence of ICP0 mRNA. The question we posed in this study is whether LAT affects the accumulation of the antisense ICP0 mRNA through complementary base-pairing.
HSV DNA consists of two covalently linked, unequal size stretches of unique sequences designated UL and US, respectively. Each sequence is flanked by inverted repeats. The inverted repeats of the L component are designated as ab and b'a' whereas those of the S component are designated as c'a' and ca. The circular DNA thus has two L-S junctions represented as baac and c'a'a'b' (Roizman 1979). The gene encoding ICP0 maps in the c sequence and presents in two copies per genome. The sequence encoding LAT-P traverses the baac junction and terminates 2235 bp upstream from the termination of transcription of ICP0 encoding gene.
The 2.0-kb LAT intron is complementary to, and partly overlaps ICP0 transcript. It has been suggested that the stable LAT intron might function as an antisense repressor of ICP0 expression (Ho and Mocarski 1989). By stably expressing the 2.0-kb LAT intron in 293 cells, Burton et al. reported the LAT affected ICP0 mRNA expression from the viral genome instead of involving a direct-acting antiICP0 antisense mechanism (Burton et al. 2003). Chen et al. reported that LAT promoter with the removel of LAT and 5' transcribed sequence does not increase ICP0 transcripts in latently infected mouse ganglia (Chen et al. 2002). However, that LAT promoter deletion mutation resulted in decreased lytic gene transcription during latency has been described in another study (Nicoll et al. 2016). Here we constructed a mutant virus by inserting two poly(A) sequences derived from SV40 and BGH viruses into LAT at its particular region to early terminate LAT transcript without touching the complementary sequence of ICP0 transcript. The purpose of this design is not to delete any region of LAT or ICP0 in HSV-1 genome, but to investigate the roles of LAT-ICP0 anti-sense binding in affecting ICP0 mRNA accumulation.
-
HEp-2, HEK293T and Vero cells were obtained from American Type Culture Collection (ATCC). The protocols for maintenance of the cells were described elsewhere (Huang et al. 2019).
-
The protocol of HSV-1 BAC constructions were previously reported elsewhere (Yan et al. 2019). Recombinant viruses SR1601 and SR1603 used in this study were constructed as follows: BAC1601 was constructed by replacing the L-S junction comprising the sequences b'a'a'c' in HSV-1(F) BAC with the sequences encoding the green fluorescent protein driven by the CMV immediate early promoter. BAC1603 was constructed by insertion of a 573 nt DNA fragment comprising the poly(A) signals of SV40 and BGH at the at two HpaI restriction enzyme sites of LAT intron from 5857 to 6027 nt in order to terminate the transcription of LAT.
Plasmid DNAs isolated from BAC1601 and BAC1603 were transfected into Vero cells. Plaques obtained on Vero cells were purified three times on Vero cells. Recombinant viruses SR1601 and SR1603 were harvested from individual plaques and viral DNA was isolated and analyzed by PCR and sequencing to verify that no adventitious sequence was introduced. Constructions of recombinant virus on the basis of BAC system would spontaneously inactivate thymidine kinase genes. The tk gene was rescued by co-transfection of the plasmid encoding tk with recombinant BAC plasmid DNA (Sears 1985).
-
Five-week-old female Balb/c mice were purchased from Charles River Laboratories (Beijing, China) and housed under specific pathogen-free (SPF) conditions. Mice handling conformed to the requirement of National Institute of Health Guide for the Care and Use of Lab animals. 107 PFU of recombinant viruses SR1601 or SR1603 were inoculated in mice through the corneal route as previously reported (Du et al. 2011). All institutional and national guidelines for the care and use of laboratory animals were followed.
-
Cells were harvested at indicated time points and lysed with RPIA lysis buffer (Beyotime) supplemented with 1 mmol/L protease inhibitor phenylmenthylsulfonyl fluoride (PMSF) (Beyotime). Cell lysates were denatured and separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), then transferred to polyvinylidene difluoride (PVDF) membranes (Millipore). The target proteins were detected by incubating with appropriate primary antibodies and horseradish peroxidaseconjugated secondary antibody (Pierce) step by step, followed by developing with the ECL reagent (Pierce) and exposed to a film. Antibodies against ICP0, GAPDH have been described in previous studies (Huang et al. 2019).
-
Total RNA was isolated by using TRI Reagent solution (Invitrogen) with DNase I (Takara) treatment. RNAs of mice trigeminal ganglia were extracted with mirVana miRNA isolation Kit (Ambion) according to the manufacturer's instructions. cDNA was synthesized from 0.5 μg total RNA by using the ReverTra Ace qPCR RT kit (Toyobo) according to the manufacturer's protocol. miRNA was reverse-transcribed from 30 ng total RNA with specific stem-loop primer as described by using TaqMan miRNA reverse transcription kit (Applied Biosystems).
-
Expression of mRNA and miRNA were analyzed by realtime quantitative PCR by using SYBR Green Realtime PCR master mix (Toyobo) in Step one plus Real-Time PCR system (Applied Biosystems) with indicated primers which have been reported elsewhere. 18 s RNA and MAP2 were used as the normalization controls in vitro and in vivo, respectively. Relative quantity (RQ) of gene expression was determined with the 2-ΔΔCT method.
-
Virus-containing samples were harvested with 3 freezeand-thaw cycles from virus-infected cells at various indicated times post infection. Virus titer of these samples was measured by conventional plaque assay. Different tenfold dilutions of HSV-1 containing samples were incubated on monolayers of Vero cells (T25 flask) at 37 ℃, 5% CO2 for 1 h, then cell medium was aspirated, cell washed two times with PBS, and incubated with fresh medium for 3 days to allow plaque formation. Cells were fixed and stained for plaque counting. HSV-1 titer was calculated from the number of formed plaques per milliliter of sample and expressed as PFU/mL.
-
Mouse trigeminal ganglia was excised and homogenized at indicated times post virus inoculation. Total DNA was isolated by using the Qiagen Blood and Tissue DNeasy kit according to the manufacturer's protocol (Qiagen). The level of viral DNA was measured with a primer pair (forward primer: 5'-AAGGTCGGCGGGATGAG-3'; reverse primer: 5'-CGGCCGCGCGATAC-3') specific for HSV-1 tk by qPCR with SYBR Green Realtime PCR master mix (Toyobo) in Step one plus Real-Time PCR system (Applied Biosystems). Adipsin (forward primer: 5'- AGTGTGCGGGGATGCAGT-3'; reverse primer: 5'- ACGCGAGAGCCCCACGTA-3') was used as the normalization control and relative quantity of DNA was determined with the 2-ΔΔCT method.
-
Five-week-old female Balb/c mice were administered with 107 PFU of SR1601 or SR1603 viruses per cornea. Trigeminal ganglia isolated from mice latently infected with recombinant viruses SR1601 or SR1603 were incubated at 37 ℃, plus 5% CO2 in 199 V medium consisting of mixture 199 supplemented with 1% serum (medium 199 V) and 10 μg/mL rabbit antibody against nerve growth factor (Abcam) at 30 days post infection.
-
All the experiments were performed in triplicates. GraphPad Prism 8.0 was used for data analysis and the results were presented as mean ± SD. Statistical software MedCalc was used to calculate the significance value for comparison of means among various experimental groups. ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001.
Cells
Viruses
Mice
Immunoblot Analyses
RNA Isolation and Reverse-Transcription
Real-Time Quantitative PCR Analysis
Virus Titration
Ganglia DNA Isolation and Quantification of Viral DNA
Virus Reactivation
Statistical Analysis
-
In order to verify whether the interaction of the 1 kb LAT with the complementary sequences of ICP0 mRNA blocks the synthesis of ICP0, replicate cultures of HEK293T cells were co-transfected with a mixture consisting of a constant amount of plasmid encoding ICP0 pcDNA, variable (0–1.0 μg) amounts of plasmids encoding the LAT intron, and variable amounts of pcDNA to ensure that the total amount of DNA transfected into the cells was constant. At 48 h after transfection the cells were harvested and total RNA were extracted and reverse-transcribed to cDNA for further qPCR analysis of ICP0 mRNA accumulation. ICP0 mRNA levels were normalized with respect to 18S RNA and are shown as relative quantity (RQ) compared with the mRNA levels measured in the cells transfected only with ICP0 mRNA or cotransfected with pcDNA. The results shown in Fig. 1A suggest that accumulation of ICP0 mRNA was repressed by LAT-P plasmid expression. Concurrently, protein lysates from harvested cells were subjected to gel electrophoresis and immunoblotted with ICP0 antibody. As shown in Fig. 1B, accumulation of ICP0 was significantly affected by higher levels of LAT intron.
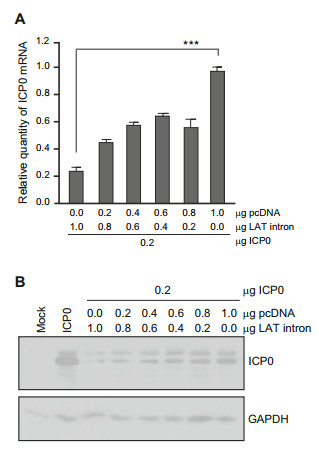
Figure 1. The effect of increased LAT plasmid expression on the accumulation of ICP0 mRNA and protein. HEK293T cells were cotransfected with ICP0, pCDNA and various amounts (0–1.0 μg) plasmids. At 48 h post transfection, cells were harvested for mRNA and protein levels detection. A Quantification of ICP0 transcript levels. Total RNA was isolated from harvested cells and analyzed by qPCR. 18S was used as internal control for normalization. mRNA levels were shown as relative quantity (RQ) compared with mRNA levels measured in the cells only with ICP0 and pCDNA cotransfection. B Western blot analysis of ICP0 protein expression. Harvested cells were lysed subjected to electrophoresis in denaturing gels and immunoblotted with ICP0 antibody. GAPDH was also blotted as a loading control. ***P < 0.001.
-
Two recombinant viruses were constructed in this study. (i)SR1601: deletion of one copy of the baac inverted repeats on the basis of HSV-1 (F); (ii) SR1603: blocked transcription of the sequences complementary to ICP0 mRNA by insertion of two poly(A) sequences designed to terminate the transcription of LAT and LAT-P.
The bacterial artificial chromosome (BAC) encoding the entire HSV-1(F) DNA sequence was described elsewhere (Horsburgh et al. 1999; Zhou et al. 2002). In the first step, we replaced the sequences of BAC encoding the internal inverted repeat region b'a'a'c' with the green florescent protein driven by the CMV early promoter (Fig. 2A and 2B). The virus derived from this BAC was designated as SR1601, which could provide technical convenience to make mutation on LAT since there is only one copy of LAT left. And the recombinant virus which replaced one copy of the repeat region with CMV-GFP has been examined for its genome stability by sequencing the whole viral genome for three generation passing from production, which has been previously described (Yan et al. 2019). Next, we replaced the sequences encoding the HpaI fragment in LAT extending from nt 5857 to 6027 with a 573 bp sequence encoding the polyadenylation signal [poly(A)] of SV40 (simian vacuolating virus 40) and BGH (bovine growth hormone) to terminate the transcription of LAT (Fig. 2C). Both poly(A) signal sequences contain the AAUAAA motif which has been reported to promote both polyadenylation and termination of transcription (Wahle 1995; Guo et al. 2011). This recombinant virus was designated SR1603. The sequences of the insertions in SR1603 are shown in Supplementary Fig. S1.
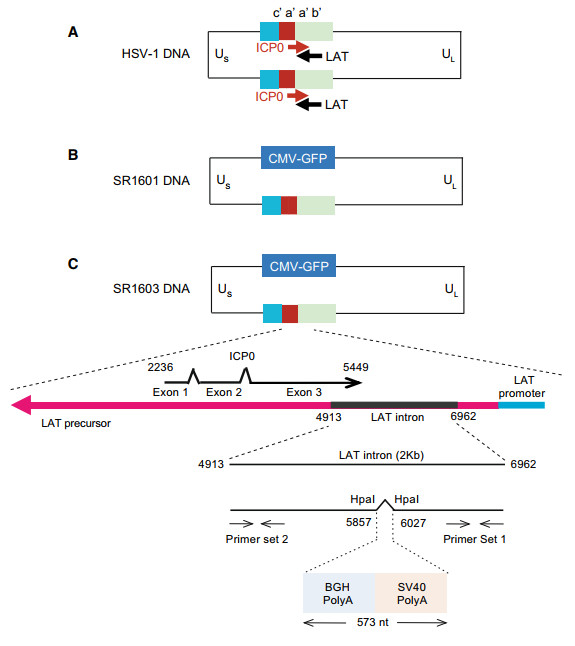
Figure 2. Construction of recombinant viruses SR1601 and SR1603. Schematic diagram of recombinant HSV-1 viruses SR1601 and SR1603 construction. SR1601 was based on parental HSV-1 F virus with IR region deletion and CMVGFP replacement. SR1603 was constructed based on BAC1601 with additional insert of SV40 polyA and BGH polyA signal sequences into LAT of TR region right before overlapping region of ICP0 gene. SV40 polyA and BGH polyA sequences were inserted into LAT of TR region for accomplishing partially early termination of LAT as indicated. Primer set 1 and set 2 designed for LAT qPCR analysis.
-
Replicate cultures of HEK293T cells were transfected with the plasmids encoding P1601 and P1603. Cells were harvested for RNA isolation at 48 h post transfection. Primer sets 1 and 2 (Supplementary Fig. S1, positions as marked in Fig. 2C) were used for examination of LAT amount. The results (Fig. 3) were as follows: Primer set 1 detected large amounts of transcripts encoding the portion of LAT upstream of the site of Insertion of the poly(A) sequences in cells transfected with both 1601 and 1603 plasmids. Primer set 2 detected large amounts of LAT sequences in cells transfected with 1601 plasmid, while that in cells transfected with the 1603 plasmid decreased to 30%. The results suggest that the poly(A) inserts partly terminated the transcription of LAT.

Figure 3. LAT accumulation in 293 T cells after transfection of the plasmids of P1601 and P1603. P1601 was constructed by insertion of 2 kb LAT intron into pcDNA3.1 and P1603 was constructed by insertion of 2 kb LAT intron carries poly(A) signals of SV40 and BGH into pcDNA3.1. 293 T cells were transfected with P1601 or P1603. At 36 h post transfection, total RNA was isolated from harvested cells and analyzed by qPCR with Primer Set1# and Set2# respectively. 18S was used as internal control for normalization. Data are mean ± SD from three independent experiments, *P < 0.05.
-
Replicate cultures of HEp-2 cells were exposed to 0.1 PFU of SR1601 or SR1603 per cell. The inoculum was replaced with fresh medium at 2 h after incubation. At 3, 6, 18 or 48 h post infection, the progeny virus was harvested and tittered in Vero cells. As shown in Fig. 4A, no significant differences of virus yields observed between SR1601 and SR1603-infected HEp-2 cells.
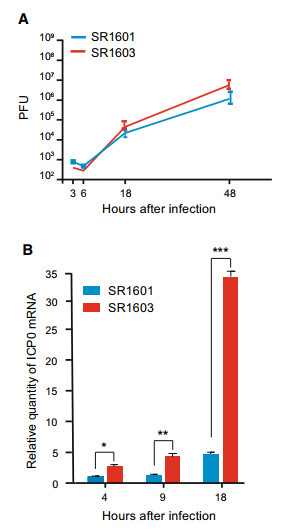
Figure 4. Characteristics of recombinant viruses SR1601 and SR1603. A Quantification of viral yields. HEp-2 cells were exposed to 0.01 PFU of SR1601 or SR1603 viruses. 2 h post infection, the inoculum was replaced with fresh medium. The virus progeny was harvested at indicated time and titered in Vero cells. B Quantification of ICP0 mRNA. HEp-2 cells were exposed to 5 MOI of SR1601 or SR1603 viruses. Total RNA was isolated from harvested cells at indicated time post infection and analyzed by qPCR. 18S was used as internal control for normalization. Data are mean ± SD from three independent experiments. *P < 0.05; **P < 0.01; ***P < 0.001.
Replicate cultures of HEp-2 cells were exposed to 5 PFU of SR1601 or SR1603 viruses per cell. The inocula were replaced with fresh medium at 2 h after infection. At times indicated in Fig. 4B, the cells were harvested and the RNAs were extracted and analyzed for the accumulation of ICP0 mRNA. The results shown in Fig. 4B suggest that SR1603 ICP0 mRNA levels were two to three fold higher at 4 and 9 h after infection and nearly eightfold higher at 18 h post infection.
-
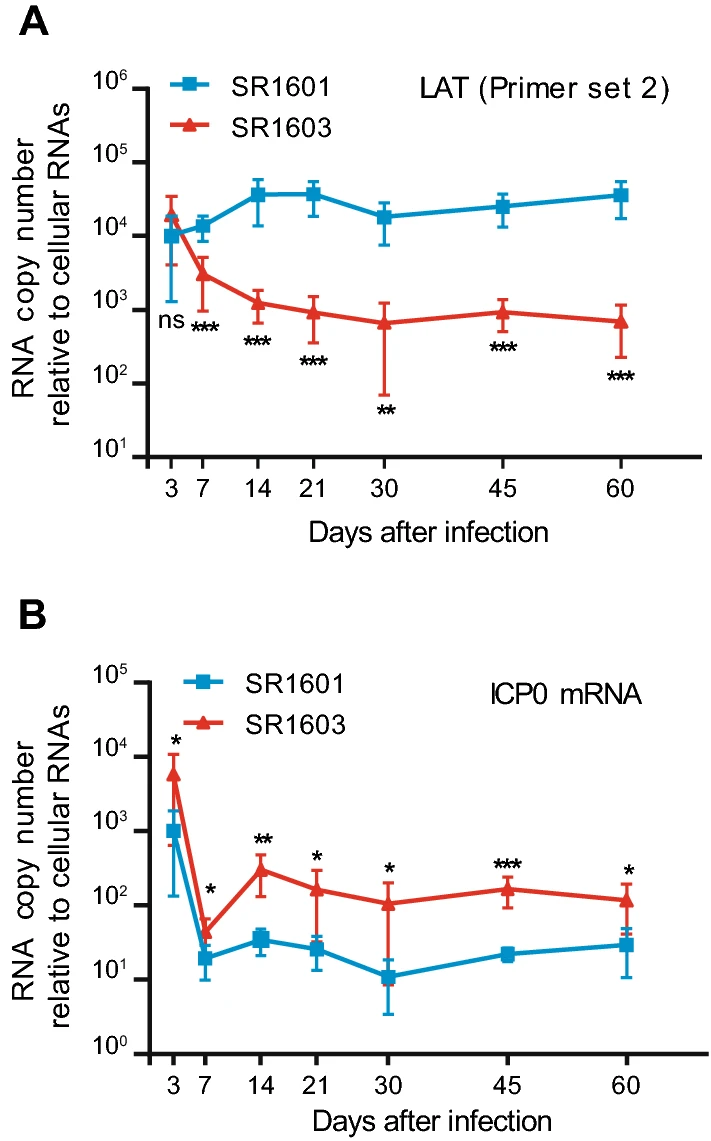
In this series of experiments, 5-week-old female Balb/c mice were inoculated by the corneal route with 107 PFU of SR1601 or SR1603 per cornea. At 3, 7, 14, 21, 30, 45 and 60 days after inoculation, mice in groups of seven were euthanized. The ganglia were excised, homogenized and tested for amounts of viral DNA as described in Materials and Methods. The results (Fig. 5) showed that the amounts of SR1601 and SR1603 DNAs increased until 21 days post infection and then slowly decreased. The amounts of DNA recovered from murine TG at day 60 approximated the amounts recovered from murine TG at day 3. As we observed, viral DNA in murine TG peaks around 14 days post inoculation. The recombinant viruses SR1601 and SR1603 basically replicate weaker than wild type HSV-1(F) in HEp-2 cells, which may result from lacking one copy of the repeat region in viral genome.
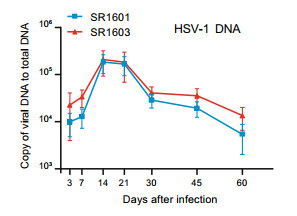
Figure 5. Viral DNA quantities of recombinant viruses SR1601 and SR1603 in latently infected murine trigeminal ganglia. 5-week-old female Balb/c mice were administered with 107 PFU of SR1601 or SR1603 viruses per cornea, 6 mice in one group. At indicated time post inoculation, mouse trigeminal ganglia were excised, homogenized. DNA was isolated and analyzed by qPCR with TK primers for examining DNA replication. Adipsin was used as internal control for normalization. Relative quantity was shown as copy of viral DNA to total DNA. Data are mean ± SD from three independent experiments.
We next examined the accumulation of the LAT and of ICP0 mRNA in SR1601 or SR1603-infected murine TG by qPCR. The levels of LAT detected in SR1603-infected murine TG were significantly lower than those accumulating in SR1601-infected murine TG starting from day 7 (Fig. 6A). In contrast, the levels of ICP0 mRNA in murine TG injected with SR1603 recombinant virus were significantly higher than those in SR1601-infected murine TG (Fig. 6B).
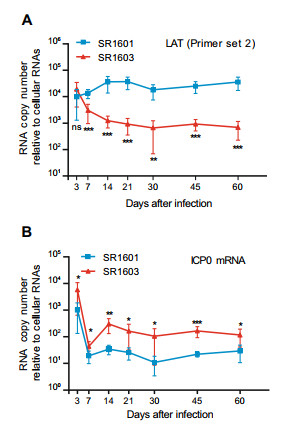
Figure 6. Accumulation of LAT and ICP0 mRNA in latently infected murine trigeminal ganglia. 5-week-old female Balb/c mice were administered with 107 PFU of SR1601 or SR1603 viruses per cornea, 6 mice in one group. At indicated time post inoculation, mouse trigeminal ganglia were excised, homogenized. Total RNA was isolated and analyzed by qPCR for examining LAT(A) by Primer set 2 and ICP0 mRNA (B) levels. MAP2 was used as internal control for normalization. mRNA levels were shown as relative quantity (RQ) compared with mRNA levels of SR1601 at 3 h post infection. Data are mean ± SD from three independent experiments. ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001.
The results in latently infected murine TG suggest that LAT antisensing to ICP0 mRNA does not have a significant impact on the amounts of viral DNA accumulation in TG harboring latent virus. The results do suggest that the amount of ICP0 expressed in infected ganglia is affected by the levels of LAT.
-
The results shown in Fig. 5 suggest that accumulation of viral DNAs in latently infected neurons is not affected by the reduced accumulation of LAT (Fig. 6A) or increased accumulations of ICP0 mRNAs (Fig. 6B). Next we wonder whether the higher levels of ICP0 affect the course of reactivation of viral genes in explanted ganglia harboring latent virus. In order to test this hypothesis, trigeminal ganglia were removed from mice at 30 days after corneal inoculation and incubated in medium 199 V supplemented with anti-nerve growth factor (NGF) antibody to accelerate the reactivation (Du et al. 2012; Zhou et al. 2013a). At 0 and 24 h post incubation, total RNA was extracted and analyzed by qPCR for the levels of LAT, miRNA-H26, and the mRNAs encoding ICP0, ICP27, TK and VP16. As shown in Fig. 7, (i) the accumulation of viral mRNAs such as ICP0, ICP27, TK and VP16, was barely detectable in neurons harboring latent SR1601 as expected, even as late as 24 h after excision and incubation of the ganglia. While the viral mRNAs in reactivated SR1603-infected murine TG were detectable starting from 0 h and increased multifold at 24 h; (ii) miRNA-H26 is synthesized very late in the course of the viral productive cycle (Du et al. 2015). The amounts of miRNA-H26 did not increase significantly after 24 h post incubation. (iii) the amounts of LAT decreased in ganglia harboring either virus at both 0 h and 24 h after excision and reactivation.
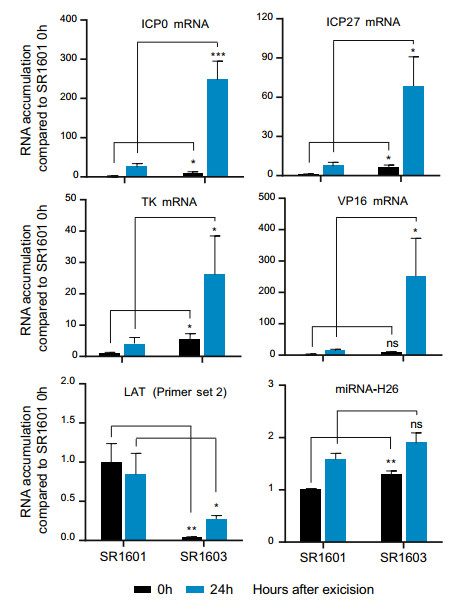
Figure 7. Expression of viral genes, miRNA and LAT after reactivation in murine trigeminal ganglia. 5-week-old female Balb/c mice were administered with 107 PFU of SR1601 or SR1603 viruses per cornea. Mouse trigeminal ganglia were removed and incubated at 37 ℃, plus 5% CO2 in 199 V medium supplemented with 10 μg/ml rabbit Ab against NGF at 30 days post infection. At 0 h and 24 h, total RNA was isolated and analyzed by qPCR for examining expression of viral genes (ICP0, ICP27, TK and VP16), miRNA-H26 and LAT by Primer set 2#. 18S was used as internal control for normalization. The expression levels were shown as relative quantity (RQ) compared with mRNA levels of SR1601 at 0 h after reactivation. Data are mean ± SD from three independent experiments. ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001.
These results indicate that a general accelerated expression of viral genes was shown in explanted ganglia harboring latent HSV with suppressed transcription of LAT.
The 2 kb LAT Intron Suppresses ICP0 mRNA and Protein Accumulation
Construction of Recombinant Viruses SR1601 and SR1603
The Insertion of poly(A) Sequences into Plasmids Encoding LAT Terminate Transcription of the DNA
Accumulation of ICP0 mRNA in the Course of Viral Replication of SR1601 and SR1603
Accumulation of Viral DNA, LAT, and ICP0 mRNA in Trigeminal Ganglia (TG) of Mice Inoculated with SR1601 and SR1603 Viruses
The Effects of Suppressed LAT Transcription on Reactivation of Latent Virus in Murine Trigeminal Ganglia
-
HSV-1 DNA entering into neurons of sensory ganglia is confronted with two alternatives: expressing the viral genes leading to productive infection or expressing LAT and suppressing the expression of genes necessary for viral replication (Zhou et al. 2013b). Precisely how the choice is made is not fully understood. Additionally, the question what prevents the activation of one or more genes in the course of latent infection is still need to be answered.
The α genes are powerful regulators of viral gene expression. They are scattered throughout the viral genome, while their position and environment do not offer clues concerning the mechanism by which they are silenced. α0, the gene encoding ICP0, is an exception. Specifically, it has been demonstrated that the abundant and stable 2.0 kb LAT intron is complementary to the 3' terminus of ICP0 mRNA (Devi-Rao et al. 1991). A longstanding hypothesis for repression of lytic gene expression during latency has been proposed: the LAT represses ICP0 expression via an anti-sense mechanism (Stevens et al. 1987; Farrell et al. 1991). If so, an increase of ICP0 transcript in latently infected ganglia would be expected when LAT expression is drastically decreased. However, a LAT deletion mutation with the LAT promoter and 5' transcribed sequences removed has been investigated that did not increase levels of ICP0 transcripts in latently infected ganglia (Chen et al. 2002). There are two uncomfortable points proposed: (i) Deletion of LAT promoter has been described that could decrease lytic gene transcription during latency (Nicoll et al. 2016). (ii) HSV-1 variant strains KOS/29 and 1704, which contain deletions of the LAT promoter region, have been found that can express a 2-kb LAT related transcript during tissue culture infection and reactivation of latently infected TG, despite LATs are not expressed during latency (Nicosia et al. 1993). Here we constructed a mutant virus by inserting two poly(A) sequences derived from SV40 and BGH viruses into LAT at its particular region to early terminate LAT transcription without touching the complementary sequence of ICP0 transcript and deleting any region of LAT in HSV-1 genome.
Subsequent studies have shown that SR1601 and SR1603 replicated to near equal levels in HEp-2 cells infected and maintained under identical conditions. Besides, viral DNA levels accumulating in sensory ganglia over a 60-day period were nearly identical. On the other hand, less LAT and increased amounts of ICP0 mRNA were examined in neurons of ganglia harboring latent SR1603 compared to neurons harboring latent SR1601 virus as expected. The results suggest that transcription of LAT-P and LAT intron indeed affect the synthesis and accumulation of ICP0 mRNA, while increased synthesis of ICP0 did not affect the accumulation of viral DNA in neurons harboring latent virus.
As noted above, viral genes reactivated and expressed in neurons of sensory ganglia excised and incubated in appropriate medium. Studies presented here show that the rate of reactivation of genes encoded by SR1603 virus was significantly higher than that of genes encoded by SR1601 virus. The results suggest that ICP0 may act as an activator of viral protein synthesis in permissive cells.
In summary, the overlap of ICP0 and LAT-transcribed regions could downregulate the accumulation of ICP0. In the absence of the overlap, an increase was observed in the transcript levels of viral genes in reactivating virus but no effect on the accumulation of viral DNA in latently infected neurons.
-
These studies were supported by grants from Shenzhen Overseas High-Caliber Peacock Foundation KQTD2015071414385495, Shenzhen Science and Innovation Commission Project Grants JCYJ20180306173333907 to Shenzhen International Institute for Biomedical Research.
-
HJ, JW, XL, GZ, WF designed research; HJ, JW, XL, RL, MZ, MC, YL performed research; HJ, JW, GZ, WF analyzed data, GZ, WF wrote and finalized the paper. All authors read and approved the final version of the manuscript.
-
The authors declare no conflict of interests.
-
All animal studies were done in accordance with guidelines and protocols approved by the Institutional Animal Care and Use Committee of the Shenzhen International Institute for Biomedical Research.


 DownLoad:
DownLoad: