







2010年25卷1期
|

作为HSV-1即早期蛋白之一,在感染细胞中ICP22是一个定位于细胞核的多功能病毒调节蛋白。在实验动物模型和一些非人类细胞系中,ICP22蛋白是必须的。但是在Vero或HEp-2细胞中是非必须的。ICP22被病毒和细胞激酶广泛地磷酸化,被酪蛋白激酶II核苷酸化。有研究表明ICP22对有效的早期蛋白还有一部分晚期蛋白的有效表达是必须的。ICP22和UL13激酶一起介导了RNA聚合酶II的磷酸化。ICP22和UL13激酶还一起介导了cdc2的激活,周期蛋白A和周期蛋白B的降解以及获得一个新的cdc2相互作用蛋白, 即UL42 DNA聚合酶加工因子。cdc2–UL42复合体介导了依赖ICP22途径的拓扑异构酶IIα的转录后修饰以促进晚期基因的表达。另外ICP22和cdk9以一个依赖Us3激酶的方式磷酸化RNA聚合酶II。
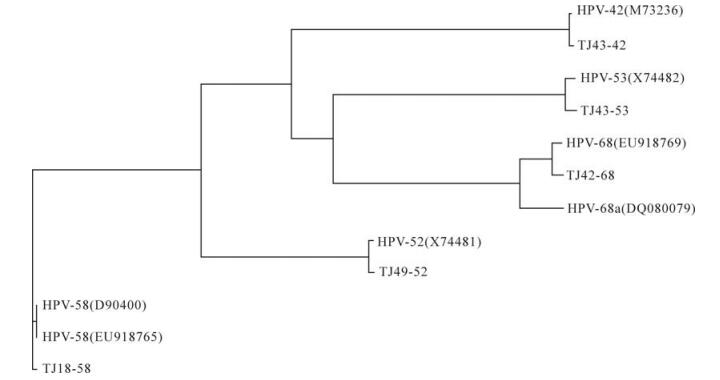
生殖器疣患者感染的HPV核酸型别及体内HPV中和抗体水平的检测
2010, 25(1): 8 doi: 10.1007/s12250-010-3078-4
本研究收集了来自天津市的生殖器疣患者的血清及棉拭子样本共50份,利用假病毒中和试验对血清中的HPV-16, -18, -58, -45, -6 和-11型中和抗体进行检测,并利用核酸分型试剂盒对棉拭子样本进行21种核酸型的检测。结果显示,36% (18/50)的血清样本为型特异性中和抗体阳性,中和抗体滴度范围为160-2560,其中22%(11/50), 12%(6/50), 10%(5/50), 4%(2/50), 4%(2/50) ,2%(1/50) 分别为HPVs -6, -16, -18, -58, -45 和 -11中和抗体阳性。而60% (30/50)的棉拭子样本为HPV DNA阳性,检出率分别为HPV-68(18%), HPV-16(14%), HPV-58(12%), HPV-33(8%),HPV-6(6%), HPV-11(6%), HPV-18(6%) 及HPV-52 (6%)。HPV DNA 与中和抗体检测结果一致性为56% (28/50),二者之间的差异具有统计学意义(P<0.05)。五种型别的(HPV -42, -52, -53, -58 和 -68)HPV全长序列被克隆并测序,结果显示这几个序列与已报道的基因组全序的同源性为98%~100%。该研究将为今后HPV感染机制研究及疫苗的研发奠定基础。
利用O型口蹄疫病毒VP1上的B细胞表位进行病毒抗体检测
2010, 25(1): 18 doi: 10.1007/s12250-010-3041-4
本研究利用编码O型口蹄疫病毒结构蛋白VP1及其氨基酸残基第141-160位和200-213位两个B细胞表位(表位1和表位2)的DNA片段,分别构建于表达载体pPROExHTb和pGEX4T-1,实现了结构蛋白VP1、GST融合表位1 (GST-E1)、串联重复的GST融合表位2 (GST-E2 (+2)) 以及GST融合的表位1-2 (GST-E1-2)在大肠杆菌胞质的可溶性表达。经Western Blot分析其反应原性后,利用纯化的VP1和上述3种融合表位建立间接ELISA方法,对80份田间猪血清检测,并与液相阻断ELISA标准方法进行对比。结果显示,建立的VP1-ELISA和三种表位ELISA与猪水泡病病毒阳性血清、豚鼠A、C、Asia1 阳性血清无交叉反应。所建立的表位ELISA和VP1-ELISA与液相阻断ELISA的相对特异性和敏感性分别为93.3% 和85.0% (GST-E1 ELISA)、95.0%、90% (GST-E2(+2) ELISA)、100% 和81.8% (GST-E1-2 ELISA)、96.6%和80.9%,该研究为进一步应用表位代替常规的细胞毒纯化抗原进行口蹄疫病毒结构蛋白抗体检测奠定了基础。
乙型肝炎病毒表面抗原适配子的筛选与鉴定
2010, 25(1): 27 doi: 10.1007/s12250-010-3091-7
存在于乙型肝炎病毒(HBV)感染细胞细胞膜上的乙型肝炎病毒表面抗原(HBsAg)为病毒感染的药物治疗提供了一个极佳的靶标,因此制备能够特异性高亲和力结合HBsAg的试剂对于HBV感染的早期诊断和治疗具有重大意义。本研究采用SELEX技术从一个含有25个随机序列的单链RNA适配子文库中筛选出了能够特异性结合HBsAg的RNA适配子,并得到一个适配子HBs-A22,该适配子能特异性结合表达HBsAg的HepG2.2.15细胞,而不结合HBsAg阴性的HepG2细胞。本研究为开发针对HBV感染细胞的靶向性诊断和治疗试剂奠定了基础。
蝙蝠SARS样冠状病毒S蛋白免疫特征分析
2010, 25(1): 36 doi: 10.1007/s12250-010-3096-2
我们前期研究中在菊头蝠里检测到了一组SARS样冠状病毒。SARS样冠状病毒与SARS冠状病毒具有一致的基因组结构和高度的序列相似性,但二者在S蛋白N端的氨基酸同源性只有64%,并且SARS样冠状病毒不能利用SARS冠状病毒的同源细胞受体(ACE2)。这些结果表明了这两组病毒又着本质的差异。为了进一步了解SARS样冠状病毒的基本特性,我们构建了表达蝙蝠SARS样冠状病毒Rp3分离株S蛋白的重组腺病毒(rAd-Rp3-S),并研究其免疫特征。结果显示重组腺病毒在小鼠体内激发了强烈的体液免疫。同时,小鼠体内IFN-γ和IL-6的水平也显著升高,进一步表明细胞免疫也产生了。然而,该重组腺病毒诱导的抗体和SARS冠状病毒S蛋白仅有弱的交叉反应,且不能中和假型SARS病毒的侵染。我们的结果不仅揭示了SARS样冠状病毒与SARS冠状病毒S蛋白免疫原性的差异,而且构建的重组腺病毒能作为候选疫苗用来防治SARS样冠状病毒的侵染。
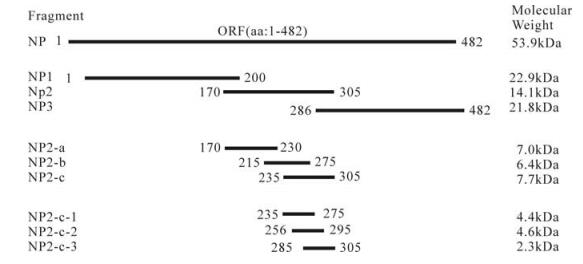
克里米亚刚果出血热病毒核衣壳蛋白的分段表达及抗原表位的鉴
2010, 25(1): 45 doi: 10.1007/s12250-010-3067-7
研究和确定克里米亚刚果出血热病毒结构蛋白的抗原特性,将有助于克里米亚刚果出血热病毒感染与免疫机制的阐明,也将为其亚单位疫苗的研制提供理论基础。本研究利用DNA重组技术将克里米亚刚果出血热病毒YL04057株S片段上核衣壳蛋白基因(NP)编码区全序列和一系列分段片段克隆入原核表达载体,在大肠杆菌中进行表达,分离纯化表达产物,用抗克里米亚刚果出血热病毒的多克隆抗体和2株单克隆抗体(14B7 和43E5)对各表达产物进行抗原表位的检测和分析。结果显示,含有NP蛋白第235-305位氨基酸的三个表达蛋白片段可以被抗克里米亚刚果出血热病毒的多克隆抗体检测出来,也可以与2株单克隆抗体杂交。表明两株单抗14B7和43E5以及兔多抗所针对的抗原表位均定位在NP蛋白的第235-305位氨基酸区域,这个区域也是NP蛋白的一个高度保守的氨基酸区域。
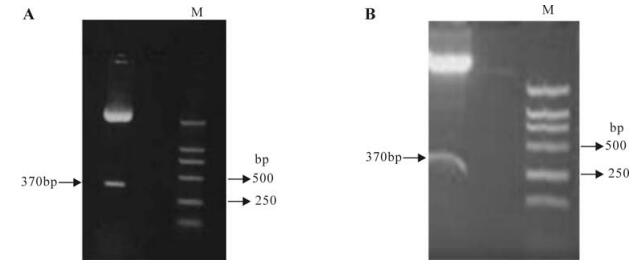
小鼠TNFR1基因的shRNA干扰质粒的构建及其在MHV-3诱导的鼠重型肝炎模型中的干预效应的初步研究*
2010, 25(1): 52 doi: 10.1007/s12250-010-3072-x
构建小鼠TNFR1(mTNFR1)的小发夹状RNA(shRNA)表达质粒p-mTNFR1shRNA,探讨其在小鼠体内对TNFR1基因表达的干预作用和规律。构建了小鼠TNFR1(mTNFR1)的小发夹状RNA(shRNA)表达质粒p-mTNFR1shRNA。利用鼠III型肝炎病毒MHV-3感染Balb/cJ小鼠制造重型肝炎动物模型;通过尾静脉高压注射将p-mTNFR1shRNA干扰质粒导入小鼠肝脏,同时注射对照质粒作为对照组,观察小鼠生存率的改变;并通过real-time PCR和免疫组化分别检测干预组和对照组的目的基因在肝脏的表达情况的变化;通过肝组织切片的HE染色观察小鼠肝脏病理改变。mTNFR1shRNA干扰质粒高压注射后,重型肝炎小鼠生存率从0提高到13.3%,并显著改善肝组织病理学变化;在基因水平和蛋白水平均可显著抑制mTNFR1在重型肝炎小鼠肝脏内的表达。成功构建了mTNFR1shRNA表达质粒p- mTNFR1shRNA,并初步证实其在体内可高效特异地抑制mTNFR1的表达,为进一步的体内干预实验奠定了基础。

猪瘟病毒RT-LAMP快速诊断方法的建立*
2010, 25(1): 59 doi: 10.1007/s12250-010-3043-2
针对CSFV的保守区设计并筛选出一套RT-LAMP引物(包括两条外引物,两条内引物以及两条环引物),经优化最终建立起简单、快速的猪瘟病毒RT-LAMP诊断方法,利用该方法可在63℃的恒温条件下一小时内大量扩增猪瘟病毒的RNA。灵敏度试验显示该方法的灵敏度比常规RT-PCR高两个数量级。特异性试验及酶切鉴定证明该方法具有很好的特异性。猪瘟病毒RT-LAMP反应可在普通水浴锅或恒温箱里完成,产物经琼脂糖凝胶电泳可见大小片段不等的斑马纹样条带;反应同时产生大量白色副产物,混浊可辨,离心后沉于管底形成白色沉淀物。猪瘟病毒RT-LAMP操作简便、快速、成本低,适合临床诊断。
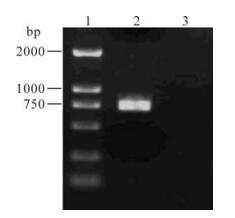
乙型流感病毒Vero细胞高产适应株的选育
2010, 25(1): 65 doi: 10.1007/s12250-010-3066-8
当流感病毒大流行的时候,用鸡胚培养方法生产的流感灭活疫苗不能满足大规模的需求。需要探索新的流感病毒培养基质。Vero细胞已作为多种人用疫苗的生产基质。所以Vero细胞可以作为生产流感病毒疫苗的细胞基质。但是大部分流感病毒株不能在Vero细胞上生长。而开发新的Vero细胞流感疫苗的前提是流感病毒株对Vero细胞敏感且保持高产的特性。通过连续在Vero细胞上传代,我们成功选育出一株乙型流感病毒Vero细胞高产适应株:B/Yunnan/2/2005va (B) 。它具备1:512的高血凝滴度,且高血凝滴度的特性一直保持到第21个代次。该毒株通过血凝抑制试验和测序两个方法进行检测。同时克隆了HA1基因,并对该基因进行分析。乙型流感病毒Vero细胞高产适应株的选育为开发新的Vero细胞流感疫苗提供了前提条件。

一种猪瘟病毒重组表位抗原间接ELISA血清学检测方法
2010, 25(1): 71 doi: 10.1007/s12250-010-3086-4
选择猪瘟病毒E2糖蛋白上包含一个抗原表位的基因区,将该基因以联接氨基酸串联,人工合成获得含有四个重复抗原表位的基因序列,并在大肠杆菌中进行表达。表达蛋白作为抗原建立一种检测猪瘟血清抗体的间接ELISA方法。检测30份阴性和80份阳性血清,计算它们的P/N值,据此得到受试者工作特征曲线(ROC曲线)。通过ROC曲线分析,确定该方法的P/N临界值为1.92。与间接血凝试验比较,该方法有97.5%的敏感性和96.7%的特异性。检测16份血清,它们的板间和板内变异系数均小于或等于6.8%。表达抗原与牛病毒性腹泻病毒的阳性血清没有交叉反应。

ISSN 1674-0769
EISSN 1995-820X
CN 42-1760/Q
主编: 石正丽
影响因子: 5.5*
*源于2022年JCR



- [01/11]《中国病毒学(英文)》期刊编辑部招聘启事
- [05/07]Q1区!VS最新影响因子5.5!
- [22/02]2022年VS高被引论文奖发布
- [21/10]第十届新生病毒性疾病控制学术研讨会 | 第一轮通知
- [09/09]肝癌细胞中CK1α上调IFNAR1的表达,从而促进I型IFN抑制HBV复制
- [09/09]一种新的干扰素诱导的长非编码RNA ZAP-IT1阻断寨卡病毒在A549细胞中的复制
- [09/09]首发精神分裂症中,驯化的人内源性逆转录病毒W家族包膜蛋白通过降低5-HT4受体的水平激活SK2
- [09/09]发热伴血小板减少综合征病毒L蛋白功能域和保守残基研究为理解病毒RNA转录/复制机制提供新思路
- [09/09]亲环素A结合AKT1并通过介导AKT/mTOR/NF-κB正反馈环路的激活促进EB病毒的致瘤作用 | VS推荐
- [09/09]转录组分析显示克里米亚刚果出血热病毒调控的关键细胞过程及III型干扰素的抗病毒作用 | VS推荐

